



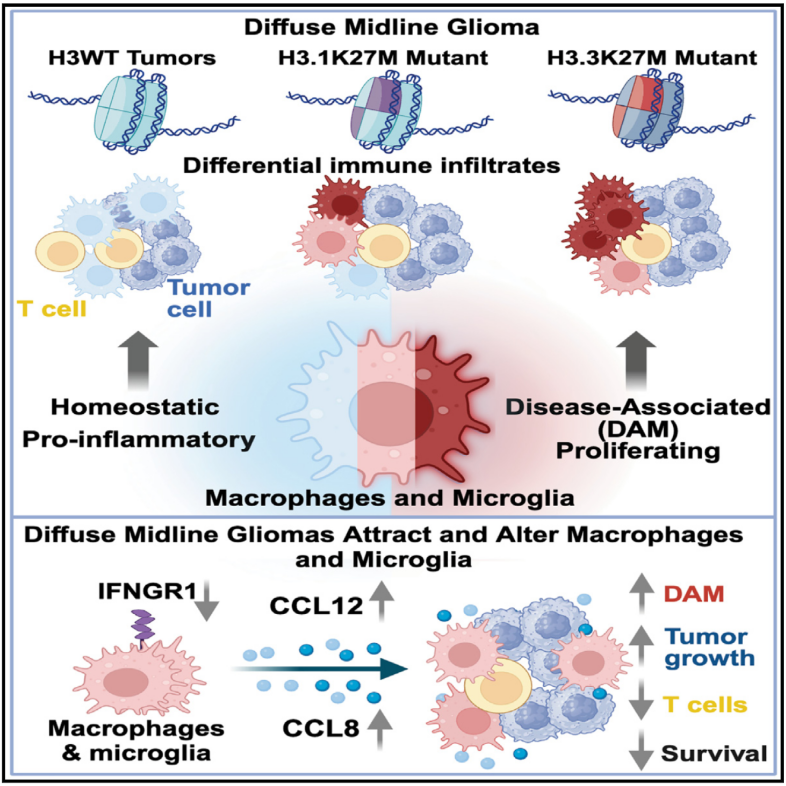
Immunity | 巨噬细胞驱动小儿高级别胶质瘤疾病发展

制版人:十一
参考文献
1. Mackay, A., Burford, A., Carvalho, D., Izquierdo, E., Fazal-Salom, J., Taylor, K.R., Bjerke, L., Clarke, M., Vinci, M., Nandhabalan, M., et al. (2017). Integrated Molecular Meta-Analysis of 1,000 Pediatric High- Grade and Diffuse Intrinsic Pontine Glioma. Cancer Cell 32, 520–537.e5. https://doi.org/10.1016/j.ccell.2017.08.017.
2. Juratli, T.A., Qin, N., Cahill, D.P., and Filbin, M.G. (2018). Molecular pathogenesis and therapeutic implications in pediatric high-grade gliomas. Pharmacol. Ther. 182, 70–79. https://doi.org/10.1016/j.pharm- thera.2017.08.006.
3. Siddaway, R., Canty, L., Pajovic, S., Milos, S., Coyaud, E., Sbergio, S.G., Vapel Anguraj, A.K., Lubanszky, E., Yun, H.Y., Portante, A., et al. (2022). Oncohistone interactome profiling uncovers contrasting oncogenic mech- anisms and identifies potential therapeutic targets in high grade glioma. Acta Neuropathol. 144, 1027–1048. https://doi.org/10.1007/s00401-022- 02489-2.
4. Wu, G., Broniscer, A., McEachron, T.A., Lu, C., Paugh, B.S., Becksfort, J., Qu, C., Ding, L., Huether, R., Parker, M., et al. (2012). Somatic histone H3 alterations in pediatric diffuse intrinsic pontine gliomas and non-brainstem glioblastomas. Nat. Genet. 44, 251–253. https://doi.org/10.1038/ng.1102.
5. Schwartzentruber, J., Korshunov, A., Liu, X.Y., Jones, D.T.W., Pfaff, E., Jacob, K., Sturm, D., Fontebasso, A.M., Quang, D.A.K., To ̈njes, M., et al. (2012). Driver mutations in histone H3.3 and chromatin remodelling genes in paediatric glioblastoma. Nature 482, 226–231. https://doi.org/ 10.1038/nature10833.
6. Larson, J.D., Kasper, L.H., Paugh, B.S., Jin, H., Wu, G., Kwon, C.H., Fan, Y., Shaw, T.I., Silveira, A.B., Qu, C., et al. (2019). Histone H3.3 K27M Accelerates Spontaneous Brainstem Glioma and Drives Restricted Changes in Bivalent Gene Expression. Cancer Cell 35, 140–155.e7. https://doi.org/10.1016/j.ccell.2018.11.01 5.
链接:http://www.lewenyixue.com/2024/10/23/Immunity%20%7C%20%E5%B7%A8%E5%99%AC%E7%BB%86%E8%83%9E%E9%A9%B1%E5%8A%A8%E5%B0%8F%E5%84%BF%E9%AB%98/

赶快来坐沙发