Science揭示人类癌症B细胞图谱
肿瘤浸润 B 细胞在肿瘤免疫中发挥重要作用, 能够作为免疫治疗应答的预测因子。因此, 分析不同癌症中 B 细胞的丰度和分化状态 可能改善免疫治疗反应。 小编今天和大家分享一篇今年 五月 刚刚发表在 SCIENCE(IF: 56.9) 杂志上全面分析人类癌症浸润 B 细胞的文章。该 文章绘制了人类癌症浸润 B 细胞 (TIBs) 的图谱,揭示了 TIBs 的不同发育模式及其对肿瘤免疫的调控作用,为基于 B 细胞开发肿瘤免疫治疗策略提供了重要基础 。

文章全面分析了 20 种癌症中 269 例患者的肿瘤、淋巴结转移灶、邻近正常组织和外周血样本的 scRNA-seq 数据 。研究首先整合了 B 细胞受体 (BCR) 测序和基因表达数据,对 B 细胞和抗体分泌细胞 (ASCs) 间的动态转化进行了详细刻画。接着研究基于不同癌症 B 细胞的单细胞染色质可及性数据 (scATAC-seq) ,分析了调控 B 细胞发育的表观网络。此外,研究也将 B 细胞定位在成熟和未成熟的三级淋巴结构 (TLSs) 中,并研究 B 细胞特异性反应的潜在调控因子。研究发现 B 细胞和浆细胞具有显著异质性,并识别出 15 个 B 细胞亚群和 10 个浆细胞亚群。研究也计算并验证了经典生发中心 (GC) 和替代的滤泡外 (EF) 两种独立的具有癌症类型偏好的 ASCs 发育路径。最后研究证实 EF 为主的癌症中非典型记忆 (AtM)B 细胞与免疫应答失调和较差的临床结局相关。

1. 人类癌症 浸润 B 细胞 的 转录 组 图谱及多样性
文章首先总体刻画了人类癌症浸润 B 细胞的转录图谱,并分析了 B 细胞转录多样性 。文章筛选了 TCGA 中 B 细胞评分较高的癌症进行采样,从 66 例患者的 153 份肿瘤、淋巴结转移、癌旁和外周血样本中分选了 CD19 + B 细胞,并进行配对单细胞 RNA 测序和单细胞 B 细胞受体测序 (scBCR-se q ,图 1A) 。整合其他已发表的公共数据后,研究最终建立了来自 20 种癌症的 269 名患者的 477 个样本的单细胞转录图谱。经过质量控后,研究共纳入 474,718 个 B 细胞的转录数据,以及来自 15 种癌症的 61 名供者的共计 166,733 个细胞的 scBCR-seq 数据。研究接着识别了 15 个 B 细胞亚群 ( 图 1B) ,其中包括 1 个 naïve B 细胞, 3 个活化 B 细胞 (ACBs ) 、 1 个 Bm 、 3 个 GC B 细胞和 2 个 ASCs( 图 1C) 。接下来研究观察到 TIBs 比非肿瘤浸润 B 细胞含有更多的 IGHG 和 IGHA ,而血液中 B 细胞富含 IGHD 和 IGHM( 图 1D) 。研究通过比值比 (OR) 分析也观察到 HSP + B 细胞、 EGR1 + ACBs 和 浆母细胞 ( PBs ) 表现出强烈的肿瘤倾向性; MT1X + B 细胞、循环 GC B 细胞、前 GC B 细胞、 GCLZ 、 GCDZ 和 IFIT3 + B 细胞在淋巴结转移中显著富集; TCL1A + naïve B 细胞和 DUSP4 + AtM B 细胞在血液中最高; MZB1 + 浆细胞 ( PCs ) 在癌旁组织中占优势 ( 图 1E) 。 TIB s 也被观察到具有显著更多的 IGH 突变, GC B 细胞和两个 ASC 亚群也表现出肿瘤特异性克隆扩增 ( 图 1F) 。
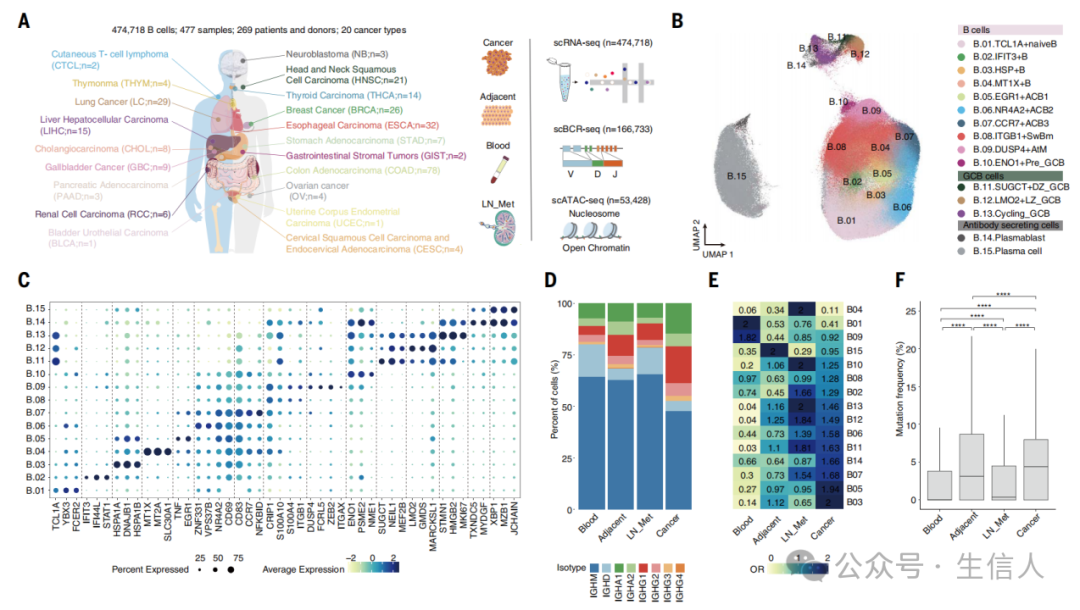
文章接着对终末分化 B 细胞 ASCs 的异质性进行了分析 。研究首先在 ASC 区域识别了 10 个具有特定基因特征和组织分布的 PC 亚群 ( 图 2A 和 B) 。其中, PC04 、 PC0 5 和 PC0 8 在肿瘤中占优势 , PC09 和 PC10 在癌旁正常组织中聚集, PC02 在淋巴结转移中普遍存在; PC01 和 PC07 也在淋巴结转移和血液中富集程度 相似; PC03 在在淋巴结转移和肿瘤中分布 相当, PC06 也相似地分布在肿瘤和癌旁组织中 ( 图 2C) 。研究接着基于 BCR 克隆共享和无监督聚类发现 AtM 和 Bm 细胞是两个主要的与 ASCs 共有克隆的 B 细胞,这表明 ASCs 可能起源于 肿瘤微环境 ( TME ) 中经典 GC 和替代 EF 路径 ( 图 2D) 。为了验证这一点,研究分选了来自肝细胞癌 (LIHC) 患者的肿瘤浸润的 naïve B 、 Bm 和 AtM B 细胞,并使用体外 B 细胞分化方法进行刺激,结果观察到 AtM B 细胞分化为 ASCs 的效率高于 naïve B 细胞,但低于 Bm 细胞 ( 图 2E 和 F) 。研究还发现 preGC, GC 循环 , 和 GC LZ B 细胞与 PBs 分享 BCR , IFIT3 + B 、 HSP + B 和 EGR1 + ACBs 也与 PBs共享 BCR ( 图 2G) 。此外,研究也观察到尽管 GC 和 EF 路径具有明显的患者和癌症类型偏好,但这两种路径在个体患者和癌症中均可观察到,这表明 GC 和 EF 反应是 ASCs 分化的普遍路径 ( 图 2H) 。进一步研究观察到肿瘤 EF 来源 ASCs 的 体细胞高突变 ( SHM ) 显著低于 GC 来源的 ASCs ( 图 2I) 。 EF 来源的 ASCs 也含有高干扰素诱导的免疫球蛋白 G (IgG) ,以及更高的 IGHM 富集,且在癌旁组织和血液中没有发生 转换重组 ( CSR ,图 2J) 。 GC 来源的 ASCs 也在肿瘤、癌旁和血液中表现出显著较高的扩增和 CSR( 图 2K) 。研究推断分化状态后观察到 GC 来源的 ASCs 倾向于处在终末分化阶段, EF 来源的 ASCs 则处于早期阶段 ( 图 2L) 。研究也揭示了 EF 和 GC 来源 ASCs 间的 BCR 库和转录特征的差异,突出了它们在抗肿瘤反应中的独特作用 ( 图 2M) 。
 文章接着分析了调控 EF 和 CG 路径的因素 。研究基于配对的 scRNA-seq 和 scATAC-seq 数据分析 TIBs ,结果观察到 8 个 B 细胞亚群,包括 naïve 、 ACBs 、 Bm 、 AtM 、 GC 和 PC 亚群 ( 图 3A-C ) 。研究通过 分析 TF 也确定了 B 细胞亚群间共享和特异的调控模式 ( 图 3D-F ) 。 这些结果揭示了一个动态的表观调控网络,该网络在癌症生态系统中能够对 B 细胞的分化和选择进行微调 ( 图 3G) ,这些结果也证明了 TF 在调控 EF 和 GC 路径平衡方面的基本作用。
文章接着分析了调控 EF 和 CG 路径的因素 。研究基于配对的 scRNA-seq 和 scATAC-seq 数据分析 TIBs ,结果观察到 8 个 B 细胞亚群,包括 naïve 、 ACBs 、 Bm 、 AtM 、 GC 和 PC 亚群 ( 图 3A-C ) 。研究通过 分析 TF 也确定了 B 细胞亚群间共享和特异的调控模式 ( 图 3D-F ) 。 这些结果揭示了一个动态的表观调控网络,该网络在癌症生态系统中能够对 B 细胞的分化和选择进行微调 ( 图 3G) ,这些结果也证明了 TF 在调控 EF 和 GC 路径平衡方面的基本作用。
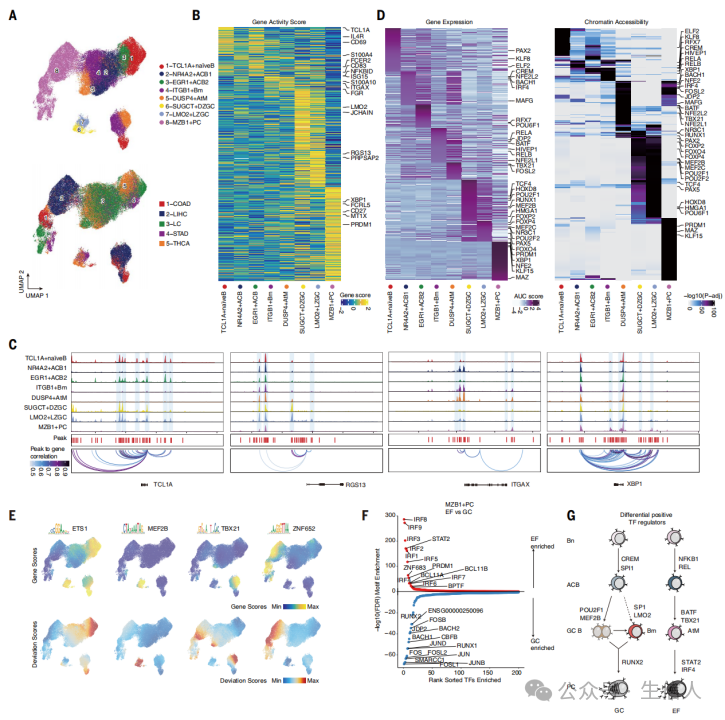
图3 人类癌症中EF和GC路径的单细胞表观基因组图谱
文章接着分析了 EF 路径主要祖细胞 AtM B 细胞的表型和功能 。研究首先观察到肿瘤浸润的 AtM B 细胞中 CD27 、 CD38 、 IRF8 和 SDC1 的表达降低,同时与 Bm 细胞 相比 PRDM1 、 IRF4 和 XBP1的表达增加, XBP1 是已知的能够抑制 B 细胞增殖并促进向 ASCs 分化的基因,这些结果表明 AtM 具有向 ASCs 分化的潜能 ( 图 4A) 。此外,研究也观察到 AtM B 细胞也高表达 BCR 信号 (SYK) 、免疫调节和激活基因 ( 图 4B) 。研究也观察到来自 AtM B 细胞的 ASCs 能够产生较高水平的自身抗体 ( 图 4C) 。接下来研究通过伪时序轨迹分析也发现 AtM 和 GC B 细胞在两个独立的分支上完全分离,起始分支为 naïve B 细胞,末端分支为 AtM( 路径 1) 和循环 GC B 细胞 ( 路径 2)( 图 4D) 。沿着这一轨迹,高亲和力特征评分和 CD24 、 CD38 、 SELL 在路径 2 中逐渐上调,而低亲和力和耗竭相关特征评分相关基因在路径 1 中相应上调 ( 图 4E) 。总的来说,这些结果表明 AtM 和 GC B 细胞经历了独立的发育过程,进而衍生出 EF 和 GC ASCs 。
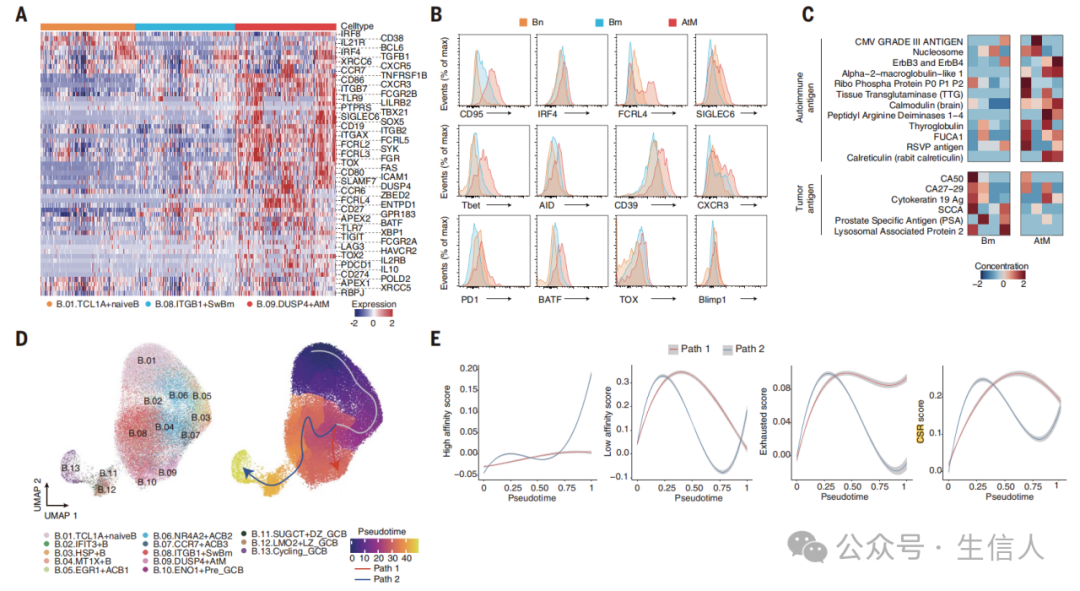
文章接着分析了三级淋巴结构 ( TLSs ) 与 TIBs 尤其是 AtM B 细胞间的空间结构和关系 。文章首先筛选了来自 LIHC 和 COAD 不同 TLS 的全组织切片,接着根据 T 和 B 细胞的细胞数量以及 fDC 染色将 TLSs 分为四个阶段 ( 图 5A) ,结果观察到 AtM B 细胞显著富集于未成熟的 TLSs ,而 Bm 主要位于成熟的 TLSs( 图 5B 和 C) 。研究接着在空间上计算了 AtM B 细胞在 TLSs 或滤泡内外的密度以及 AtM B 细胞到界面的距离,结果观察到 AtM B 细胞主要位于未成熟 TLSs 的中心 ( 图 5D 和 E) 。此外,研究也观察到 AtM B 细胞与 PD1 Hi 耗竭 T 细胞呈正相关 ( 图 5F) 。研究通过配体受体分析细胞互作也观察到 PD1 Hi CD4 + Tph 细胞和 AtM B 细胞间存在交互,这一结果表明 PD1 Hi CD4 + Tph 细胞在促进 AtM B 细胞分化方面具有潜在作用 ( 图 5G) 。研究通过分析已发表的空间转录组学数据集也观察到 CD4 + PD1 Hi Tph 和 AtM B 细胞在 TLSs 中呈正相关 ( 图 5H 和 I) 。与非 TLS 区域相比,在靠近 AtM B 细胞的 TLS 区域也观察到 CD4 + PD1 Hi Tph 细胞密度和频率显著较高 ( 图 5J 和 K) 。为了验证这一点,研究分选了 LIHC 浸润的 PD1 Hi CD4 + 外周辅助 T 细胞 ( Tph ) 、 辅助性 T 细胞 ( Th ) 和调节性 T (Treg) 细胞,并与健康供者的外周总 B 细胞或分选的 naïve B 细胞共培养 ( 图 5L) ,结果发现 Tph 细胞比 Th 和 Treg 细胞更有效地诱导 AtM B 细胞分化,而 IL-21R 阻断显著减弱了这一作用 ( 图 5M) 。总之,这些数据表明肿瘤浸润的 Tph 细胞显著促进了 AtM B 细胞的分化。

图5 AtM B细胞聚集在未成熟的TLS中由Tph通过IL-21-IL-21R轴诱导
6.谷氨酰胺代谢建立了EF AtM B细胞的表观遗传特征并影响患者临床结局
文章接着研究了机制上驱动 EF 和 GC 路径响应的因素 。研究首先 计算 Bm 和 AtM B 细胞的代谢通路评分,结果观察到 谷氨酰胺、谷氨酸和鞘脂代谢在肿瘤浸润的 AtM B 细胞中上调,而花生四烯酸代谢在 Bm 细胞中显著上调 ( 图 6A) 。因此,研究 假设谷氨酰胺代谢可能在 TME 启动 EF 反应中起潜在作用。为了验证这一假设,研究 从健康血液中收集 B 细胞,并用 R848 (TLR7 激动剂 ) 、 IFNg 和谷氨酰胺刺激这些细胞,结果观察到 谷氨酰胺显著诱导 AtM 分化,而谷氨酰胺酶 (GLS)抑制能够 显著减弱 AtM 分化 ( 图 6B 和 C) 。此外,研究也 监测了谷氨酰胺刺激下 B 细胞的氧消耗率和三磷酸腺苷 (ATP) 生成,结果观察到整体 线粒体呼吸和糖酵解减少 ( 图 6D) 。这表明谷氨酰胺可能重塑 B 细胞的代谢特征,从而使 B 细胞向 AtM B 细胞分化。接下来研究分析了 13 C 标记的谷氨酰胺经过 R848 处理后的代谢命运,结果发现 谷氨酰胺被催化生成谷氨酸,并进入三羧酸 (TCA) 循环 ( 图 6E) 。此外,研究也观察到 谷氨酰胺衍生的α - 酮戊二酸 (a-kg) 可以直接调节 AtM 分化 ( 图 6F) 。研究通过分析 ATAC-seq 和 RNAseq数据也发现 谷氨酰胺处理的 B 细胞存在稳定的 AtM 激活 ( 图 6G) 。通路富集分析也再次证明了这一结构 ( 图 6H) 。此外,研究也发现 与 Bm 和 naïve B 细胞相比,肿瘤浸润的 AtM B 细胞中 phospho-mTORC1 及其下游靶点的水平显著增加,这 表明 mTORC1 信号通路可能参与谷氨酰胺的调控 ( 图 6I 和 J) 。研究也发现 抑制 mTORC1 通路能够 显著降低谷氨酰胺刺激下的 AtM 分化 ( 图 6K 和 L) 。接下来,研究使用谷氨酰胺将外周 B 细胞诱导为 AtM ,并在体外与外周 CD3 + T 细胞共培养,结果发现 AtM B 细胞可以降低扩散 CD4 + 和 CD8 + T 细胞 ( 图 6 M ) ,并影响 T 细胞产生 IFNg 、肿瘤坏死因子的能力 ( 图 6N ) 。最后,研究分析了 Bm 和 AtM B 细胞在癌症中的预后价值,结果发现 Bm 细胞与良好预后相关,而 AtM B 细胞与较差的生存期相关 ( 图 6O) 。此外,在已发表的抗 PD 1 治疗的黑色素瘤和肺癌队列中,也观察到 AtM B 细胞的丰度与治疗耐药显著相关,而 Bm 细胞与改善的应答和更长的生存期相关 ( 图 6P 和 Q) 。总之,这些结果表明, EF 来源的 B 细胞与不成熟的 TLSs 和耗竭的 T 细胞相关,导致某些癌症的免疫治疗耐药和预后不良。
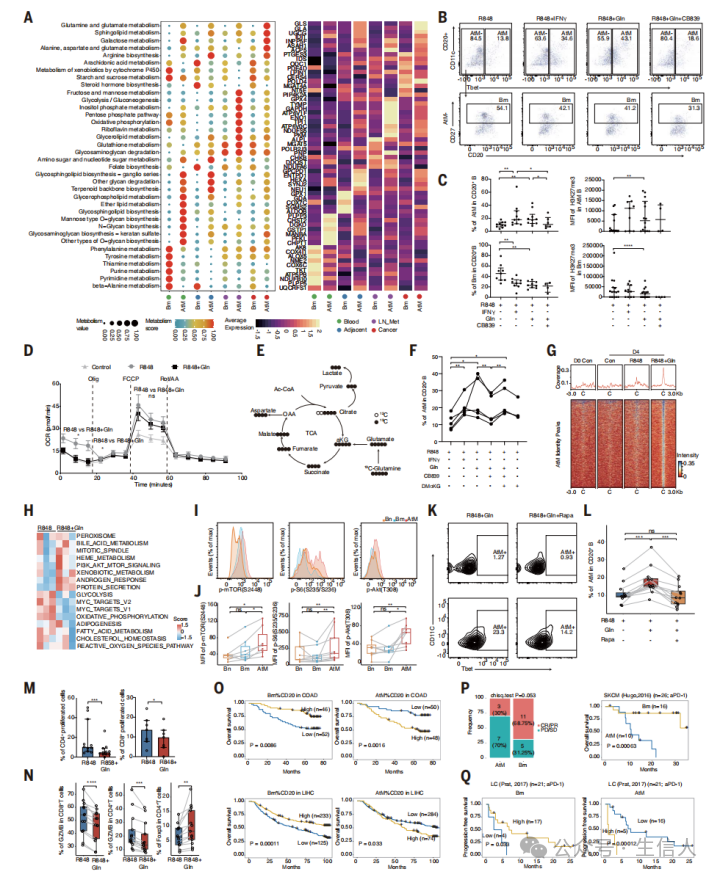 图6 谷氨酰胺促进AtM B细胞分化获得免疫调节功能
到这里文章的主要内容就介绍完了。总结一下, 该文章整合了泛癌的单细胞转录数据、 BCR 数据和基因组等多组学数据,全面绘制了人类泛癌 B 细胞图谱,并从多个角度刻画了肿瘤浸润 B 细胞的异质性、调控机制,以及动态分化路径的潜在机制 。该文章为研究癌症 B 细胞提供了重要参考,也为基于 B 细胞改善肿瘤免疫治疗提供依据及思路。
图6 谷氨酰胺促进AtM B细胞分化获得免疫调节功能
到这里文章的主要内容就介绍完了。总结一下, 该文章整合了泛癌的单细胞转录数据、 BCR 数据和基因组等多组学数据,全面绘制了人类泛癌 B 细胞图谱,并从多个角度刻画了肿瘤浸润 B 细胞的异质性、调控机制,以及动态分化路径的潜在机制 。该文章为研究癌症 B 细胞提供了重要参考,也为基于 B 细胞改善肿瘤免疫治疗提供依据及思路。
版权声明:本文为“乐问号”作者或机构在乐问医学上传并发布,仅代表该作者或机构观点,不代表乐问医学的观点或立场,不能作为个体诊疗依据,如有不适,请结合自身情况寻求医生的针对性治疗。
链接:http://www.lewenyixue.com/2024/07/01/Science%E6%8F%AD%E7%A4%BA%E4%BA%BA%E7%B1%BB%E7%99%8C%E7%97%87B%E7%BB%86%E8%83%9E%E5%9B%BE%E8%B0%B1/
THE END





赶快来坐沙发