



高分热点:肿瘤器官特异性转移的罪魁祸首—肿瘤干细胞
谈恋爱期间,一个令广大男同胞头大的问题:你到底爱我什么!为何弱水三千,只取一瓢饮? 回答不外乎为:你和她们都不一样~
同样,就肿瘤生物学行为而言, 肿瘤转移为什么偏偏要转移到特定器官,一定是被转移的器官身上有其他器官没有的特质在“吸引”着肿瘤。
肿瘤干细胞CSCs是肿瘤再生转移的罪魁祸首, 尽管肿瘤中有许多恶性细胞,但只有CSC可以驱动播散性癌症的持续和复发,并使肿瘤发生远端转移。
肿瘤转移往往表现为器官亲和性或器官特异性转移, 这可能受多种因素的调控,包括肿瘤内在因素、循环模式、器官特异性生态位以及肿瘤细胞与宿主微环境的相互作用。
原发性肿瘤中的癌细胞通过进化的适应性向特定器官趋向, 这是由表观基因组程序调节的,并通过特定基因的激活表现出来。这一过程被称为“种子预选”,这可以解释 为什么转移性倾向通常与原发性肿瘤中的特定基因表达模式相关。
让我们一起开看一下这篇IF:24.5的文章是如何探索 干细胞促进肿瘤向特定器官转移的。本研究揭示了一个 以ASCL2和PTPRO高表达为特征的干细胞亚群P1细胞通过其过表达的配体DLL4促进CAF和D内皮细胞向卵巢转移的过程。


结果
结直肠癌单细胞转录组景观
对原发性CRC、oCRC、lCRC、邻近正常组织和PBMC样本进行了 scRNA测序( 图1A )。注释出10种细胞类型,包括上皮细胞、成纤维细胞、内皮细胞和胶质细胞等4种非免疫细胞类型和B细胞、T细胞、髓细胞、肥大细胞、巨核细胞和浆B细胞等6种免疫细胞类型(图1B-C),计算了 这10种细胞类型在不同组织类型中的比例 ( 图1D-E )。

图1:人类结直肠癌的单细胞转录组景观
干细胞是结直肠癌转移的起源细胞
UMAP显示14种上皮细胞亚型(图2A) , 在14种细胞亚型中鉴定出5种恶性细胞亚型 ( 图2 B)。 肠细胞在正常结肠组织中富集,胆管细胞在lCRC中富 集, 干细胞样细胞和其他恶性细胞亚型在原发性和转移性CRC样本中均富集 (图2C)。原 发性结直肠癌和转移性结直肠癌的干细胞样恶性细胞亚型的DNA拷贝数变异明显高于邻近正常组织的上皮细胞(图2D-E)。然后,预测五种恶性细胞亚型的分化状态和14种上皮细胞亚型RNA速度 ,并对上皮细胞的干性密度进行评分,结果显示 干细胞样细胞可能是结直肠癌转移的起源细胞 。分析了 干细胞样细胞与其他细胞亚型的差异表达基因(图2H) , 在干细胞样细胞中MYC靶点V1和WNT/β-catenin信号通路中基因的显著表达升高(图2I) ,并且表达程序在恶性细胞亚型之间存在异质性。
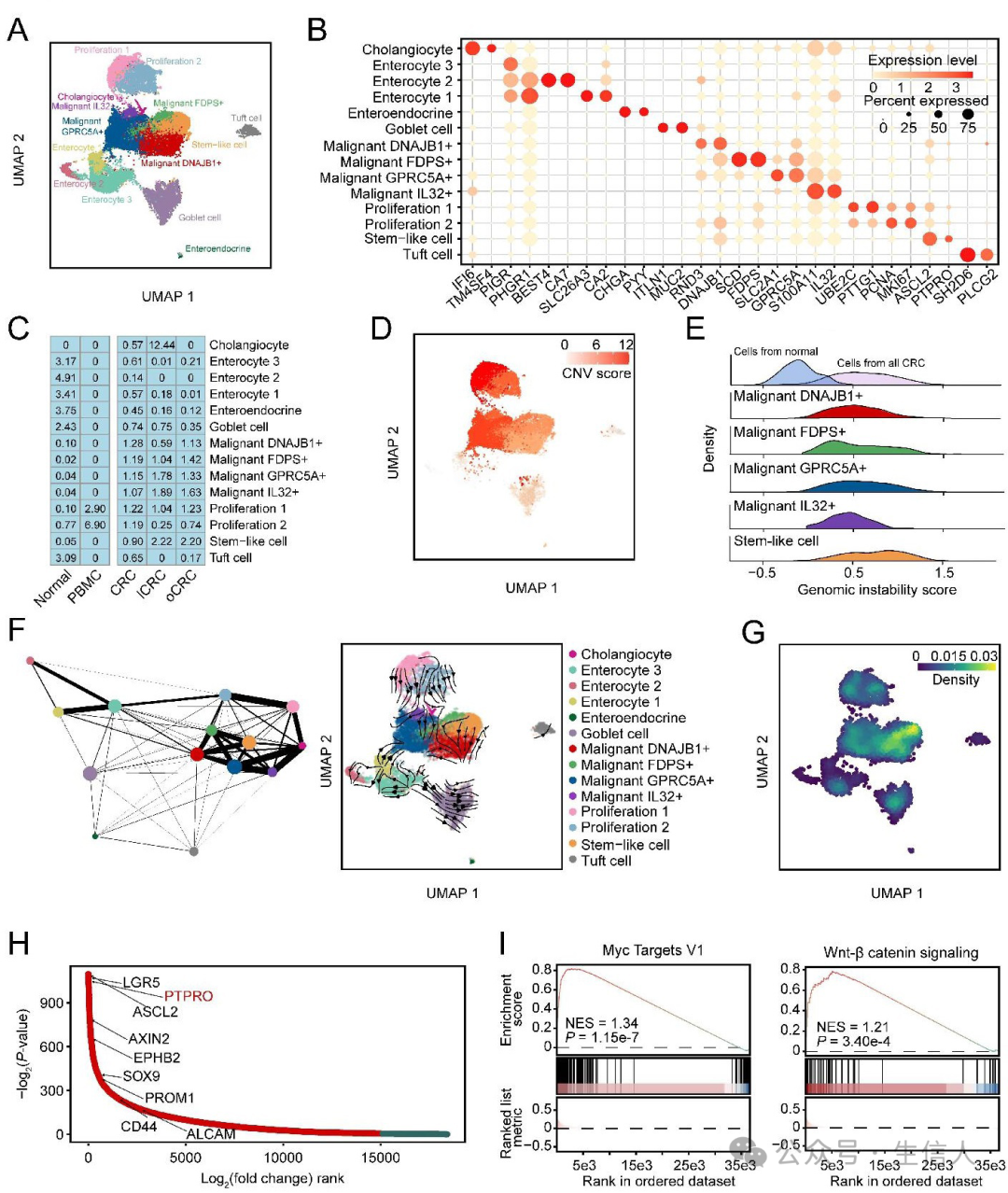
图2:干细胞是结直肠癌转移的起源细胞
ASCL2和PTPRO的过表达赋予CRC干细胞样细胞转移表型
ASCL2是肠道干细胞的关键调节因子,参与CRC的增殖和运动 , 由于本研究中CRC样本的干细胞样细胞具有高水平的PTPRO RNA(图3A),因此 分析了ASCL2和PTPRO表达水平之间的关系,并发现了显著的正相关。 于是 对原发性CRC样本进行空间转录组学和免疫荧光分析 , 发现一种亚型上皮细胞高度表达ASCL2和PTPRO(图3B-C), ASCL2可能是PTPRO的TF。 在两个CRC细胞系中进行了基因操作实验来验证这一概念 ( 图3D ), 还研究了原发性CRC肿瘤中心、侵袭性前沿和腹膜转移灶中的PTPRO RNA水平,发现 转移灶中的水平明显高于原发性肿瘤中心或侵袭性前沿 (图3E)。
通过PTPRO表达水平对MSS结直肠癌患者总生存时间的 KM估计发现 高PTPRO水平的MSS CRC患者的生存时间明显短于低PTPRO水平的患者 (图3F)。
最后检测了PTPRO表达变化对CRC细胞表型的影响,发现 PTPRO敲低显著降低了CRC细胞的迁移、侵袭(图3G)和球体繁殖能力。
总之,这些结果表明, 原发性CRC含有一种以ASCL2和PTPRO高表达为特征的干细胞亚型,这赋予了CRC转移表型。
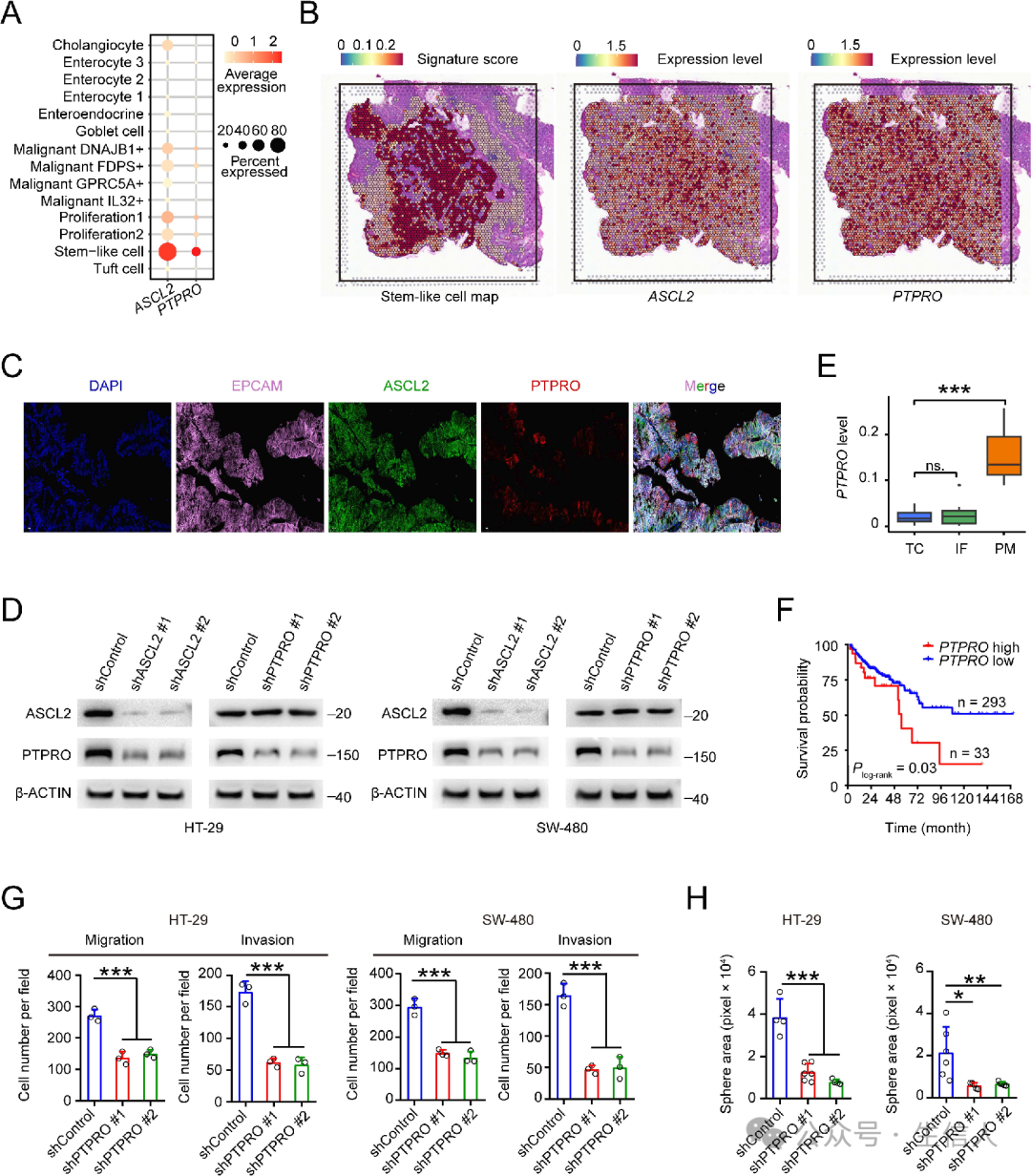
图3:ASCL2和PTPRO的过度表达赋予CRC干细胞样细胞转移表型
CRC干细胞样细胞在器官特异性CRC转移中的异质性
为了 探索干细胞亚型是否具有可能与器官特异性转移相关的异质性。基于转录组学特征,将干细胞样细胞分为8个不同的亚群(P1至P8) (图4A),比较了不同器官中干细胞亚型的分布, 正常、CRC、lCRC和oCRC中P1、P2、P3和P4细胞的比例(图4B-C) ,CRC、lCRC和oCRC细胞亚型分配及其空间分布模式( 图4 D)。
分析了 每个干细胞亚型的特征表达(图4E) ,P1和P3细胞基因表达特征的差异在原发性CRC中得到了验证(图4F),在oCRC或lCRC中保留了原发性CRC中形成的不同表达模式(图4G)。
临床组织样本免疫组化染色显示,转移至卵巢的原发性CRC 的DLL4表达水平高于转移至肝脏的原发性 CRC(图4H)。ROC的AUC 表明DLL4表达水平预测卵巢转移的能力(图4I)。
还通过尾静脉将ASCL2、PTPRO或DLL4敲低的CRC细胞移植到小鼠体内,结果显示,与对照细胞移植相比,卵巢转移率明显降低(图5A-C)。
这些结果提示, 由不同的主TF引起的干细胞样细胞的异质性可能决定了原发性结直肠癌的器官特异性转移。
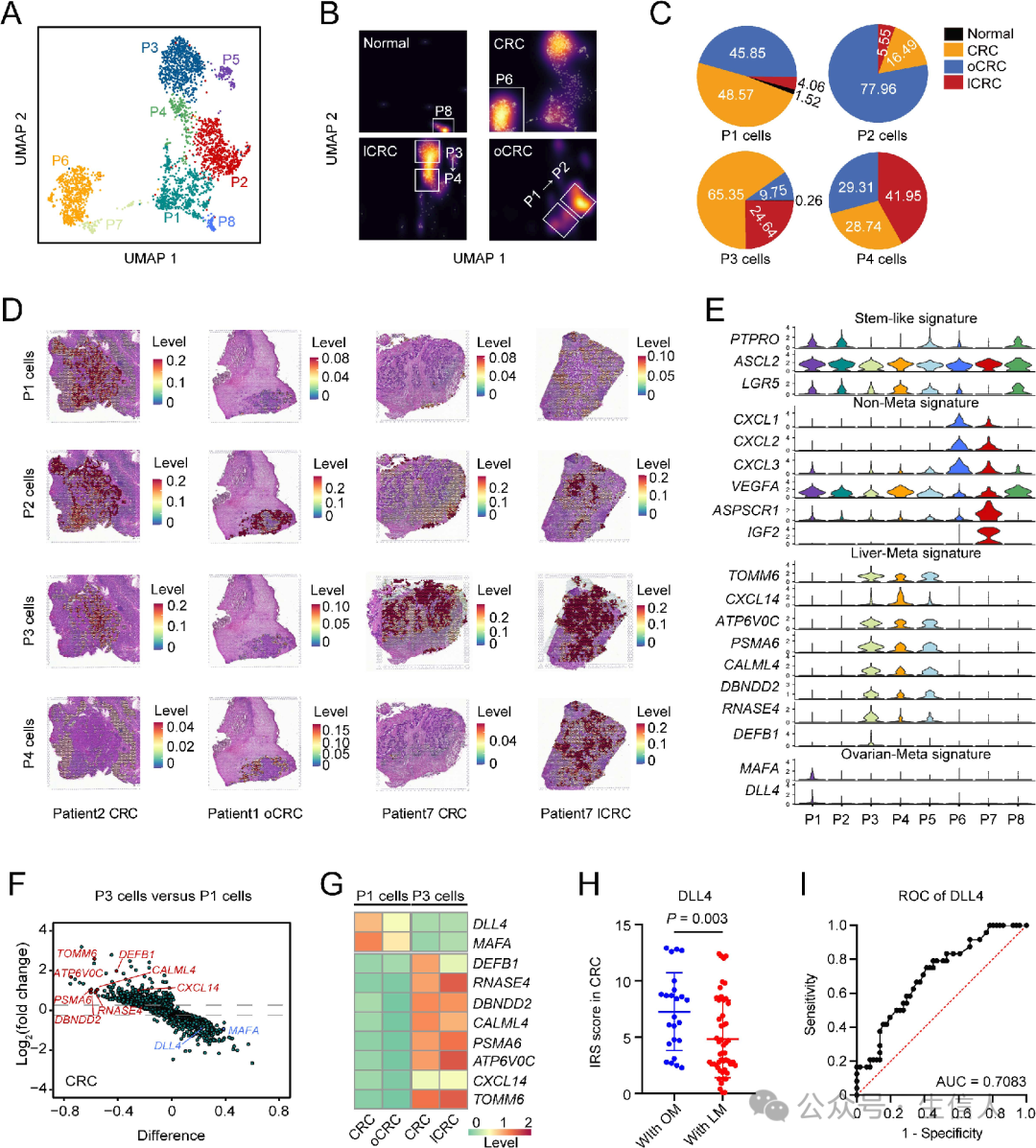
图4:CRC干细胞样细胞在器官特异性CRC转移中的异质性
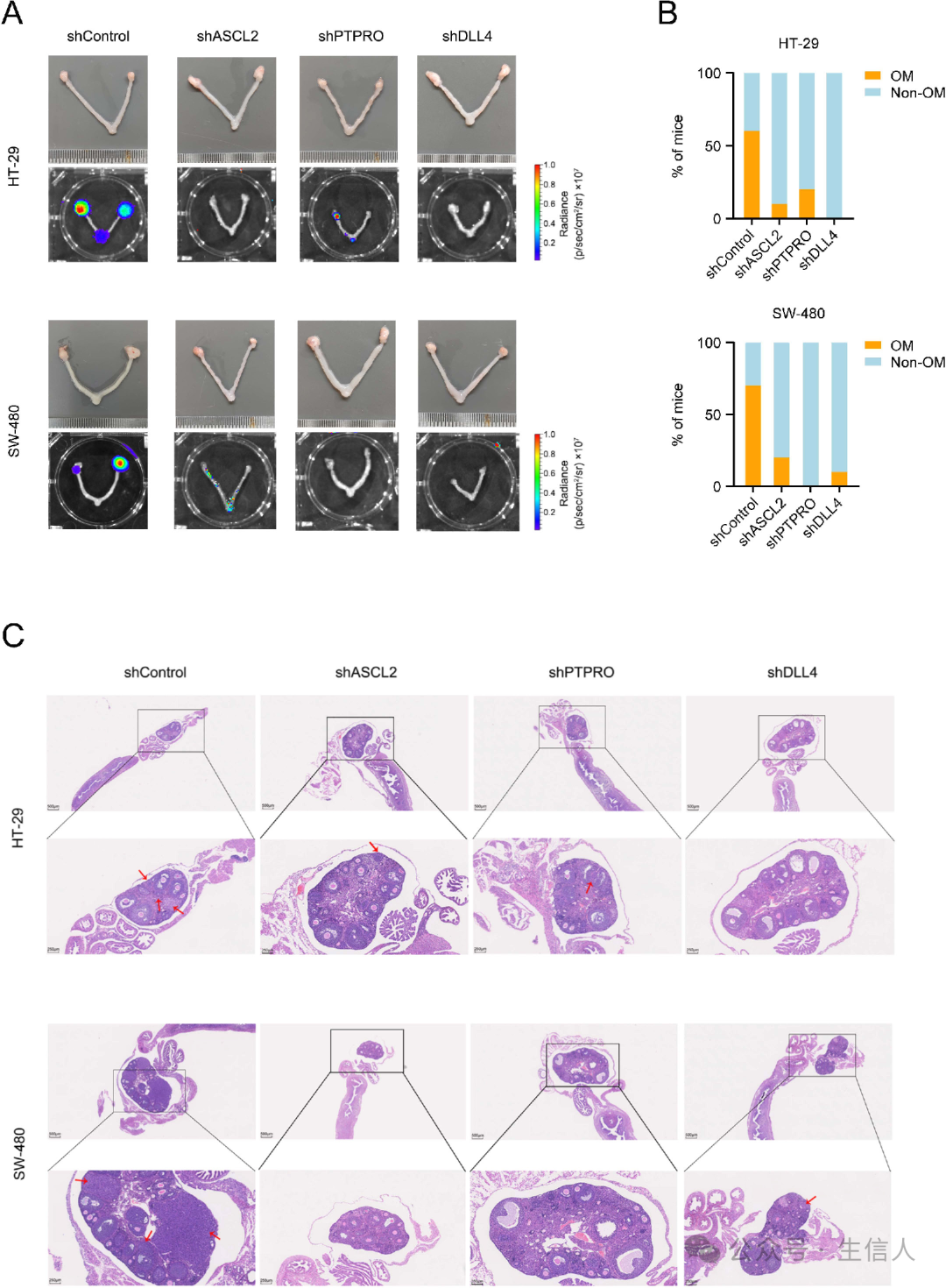
图5:DLL4、ASCL2和PTPRO促进CRC卵巢转移
干细胞样细胞与CAFs和分化内皮细胞在结直肠癌转移中的相互作用
由于癌症转移是由癌细胞自身和肿瘤微环境中其他细胞类型(包括CAFs和内皮细胞)的协同作用决定的,所以,对CAF进行了深入的研究。 在所有样本中鉴定出13种成纤维细胞亚型(图6A),计算了配体-受体对的吸引力强度, 发现P1细胞比P6细胞通过DLL4-NOTCH2、DLL4-NOTCH3和PDGFAPDGFRA的配体-受体相互作用与CAFs产生更强的串扰( 图6B ) 。基于P1-CAFs相互作用评分对癌症基因组图谱(TCGA)-COAD数据集中 MSS 结直肠癌患者生存时间的KM估计 , 评分高的MSS结直肠癌患者的生存时间明显短于评分低的患者(图6C)。
对内皮细胞进行了亚分类,重点研究了在原发性CRC中富集的内皮细胞 (图6F,6H,6I), 在原发性结直肠癌微环境中,P1细胞可以通过其过表达的配体DLL4与CAF和D内皮细胞上的NOTCH受体相互作用,从而促进CAF和D内皮细胞向卵巢转移 (图6K)。

图6:干细胞样细胞与CAFs和分化内皮细胞在结直肠癌转移中的相互作用
卵巢结直肠癌转移的代谢特征
发现了 一些oCRC微环境中富集 的细胞(图7A),对这三种肌成纤维细胞亚型进行 伪时间顺序分析(图7B) , 检查肌成纤维细胞和恶性细胞之间的代谢物传感器通信(图7C) , 在MyoFib_RBP1细胞与FDPS+恶性细胞之间的所有代谢物传感器伙伴中, l -谷氨酰胺及其转运蛋白的通讯评分明显高于其他细胞(图7D),因为FDPS+恶性细胞产生谷氨酰胺的能力较低,需要外部供应 (图7E)。
oCRC中FDPS+恶性细胞表现出氧化磷酸化、糖酵解、d -谷氨酰胺/ d -谷氨酸代谢和TCA循环异常激活(图7F),部分基因及其催化代谢产物表达水平升高(图7G-H)。人结直肠癌细胞中的GLUL mRNA水平及其培养基中的谷氨酰胺含量低于人卵巢成纤维细胞, oCRC中FDPS+恶性细胞代谢非常活跃,肌成纤维细胞可以为这些癌细胞提供谷氨酰胺进行谷氨酰胺代谢。
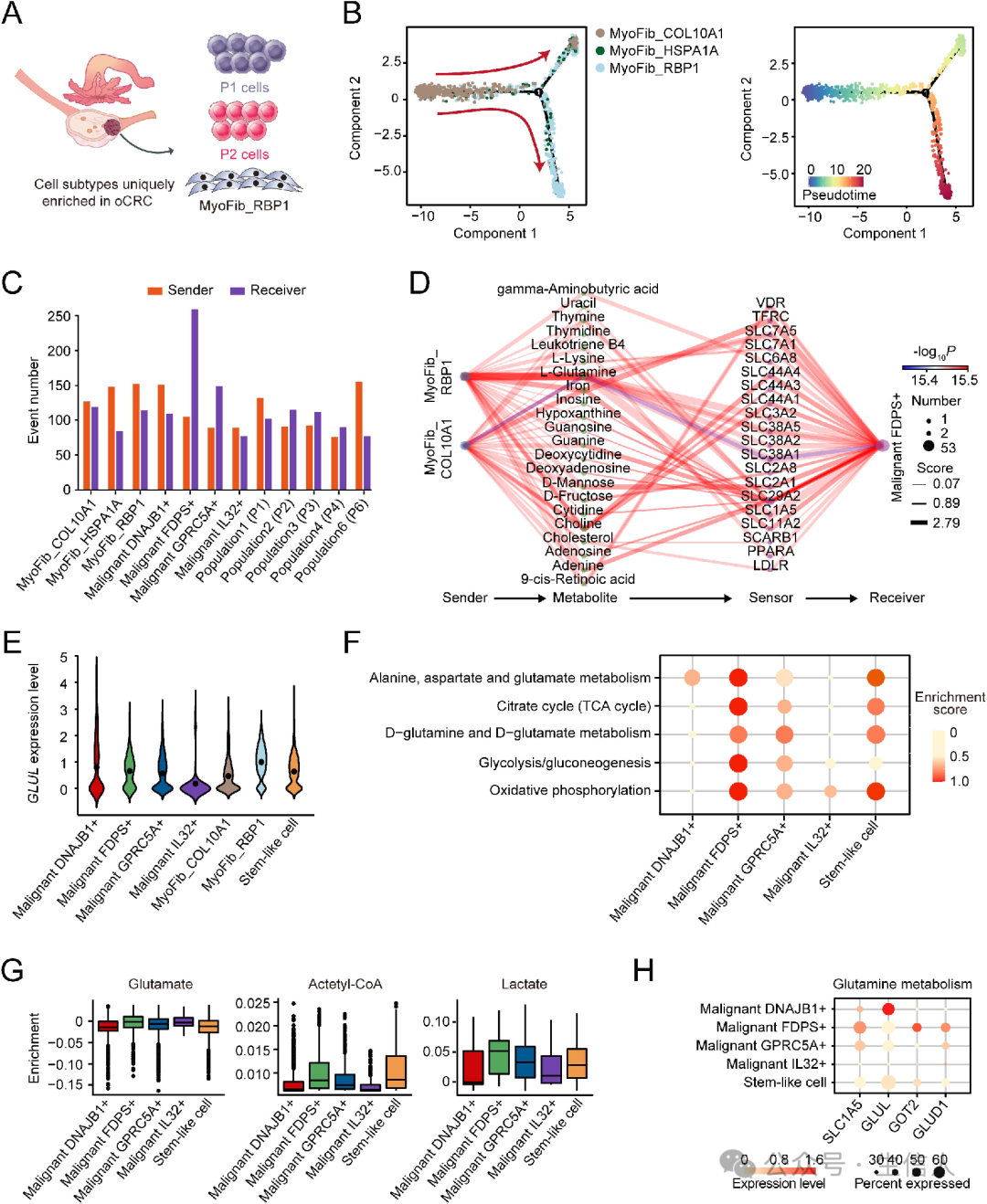
图7: 卵巢转移性结直肠癌(oCRC)代谢特征的表征
TIGIT是oCRC免疫治疗的潜在靶点
分析了oCRC免疫治疗的潜在靶点,PD-1/PD-L1对可能不是oCRC中主要的免疫检查点信号分子。相反, TIGIT-NECTIN2对可能是重要的免疫检查点分子。
通过整合scRNA-seq,空间转录组分析和功能分析, 发现了一个以PTPRO和ASCL2为转移罪魁的干细胞簇,其亚群显示出进一步不同的肝脏或卵巢器官亲和性。在原发性结直肠癌中,转移的结直肠癌细胞通过NOTCH信号通路与CAFs和D内皮细胞相互作用,获取转移所需的营养和能量。 本研究的 发现揭示了器官特异性CRC转移的罪魁祸首。
链接:http://www.lewenyixue.com/2024/08/30/%E9%AB%98%E5%88%86%E7%83%AD%E7%82%B9%EF%BC%9A%E8%82%BF%E7%98%A4%E5%99%A8%E5%AE%98%E7%89%B9%E5%BC%82%E6%80%A7%E8%BD%AC%E7%A7%BB%E7%9A%84%E7%BD%AA%E9%AD%81%E7%A5%B8%E9%A6%96%E2%80%94/
 知道|国家药品不良反应监测年度报告(2024年)
知道|国家药品不良反应监测年度报告(2024年)
赶快来坐沙发