高分思路之进阶版肿瘤干性双热点联合分析
癌症干细胞(CSCs)作为解释肿瘤异质性潜在机制的理论之一,一直备受关注,你还在单纯研究肿瘤干性吗,试一试进阶版结合双热点吧,经典的大神思路, 不仅研究肿瘤干性本身,还关注是什么影响或导致肿瘤干性,比如今天分享的这篇文章,将双热点“神经内分泌转化”与“肿瘤干性”结合,阐明了癌细胞通过去甲肾上腺素- ATF1驱动的核-线粒体协同程序获得干性。
比如今天的思路“神经免疫+肿瘤干性”,可以同样用来研究“表观遗传(组蛋白修饰、RNA修饰)+肿瘤干性”,“谱系转化(神经内分泌转化)+肿瘤干性”,“TME+肿瘤干性”,“细胞外基质+肿瘤干性”……
癌细胞接受细胞外信号输入以获得干细胞样状态,但肿瘤微环境(TME)神经信号如何引导肿瘤干细胞建立分层肿瘤结构仍然是一个谜。今天给大家分享一篇发表在《Signal Transduction and Targeted Therapy》(IF39.3)的文章,本研究对33种TCGA癌症类型的10852个样本进行了泛癌症转录组学筛选,发现 cAMP-responsive element (CRE)转录因子是癌症干细胞的趋同激活因子。单细胞转录组证实了核多能性与线粒体生物发生在癌症干细胞样细胞中的协调激活。这些发现阐明了癌细胞通过去甲肾上腺素- ATF1驱动的核-线粒体协同程序获得干性。
肿瘤干细胞是一种具有克隆性、自我更新和多谱系分化特性的功能性细胞状态。多能转录因子控制着正常干细胞的命运, TME神经信号越来越多被证明促进癌症进展。神经递质是在癌症发展过程中非常受欢迎的神经信号。在神经递质中,肾上腺素能信号诱导的去甲肾上腺素在癌症进展中起核心作用。cAMP-CREB1信号可增强癌症的发生,癌细胞是否通过使用TME神经信号依赖的CRE程序获得干细胞仍不确定,本研究就这一问题进行了研究。
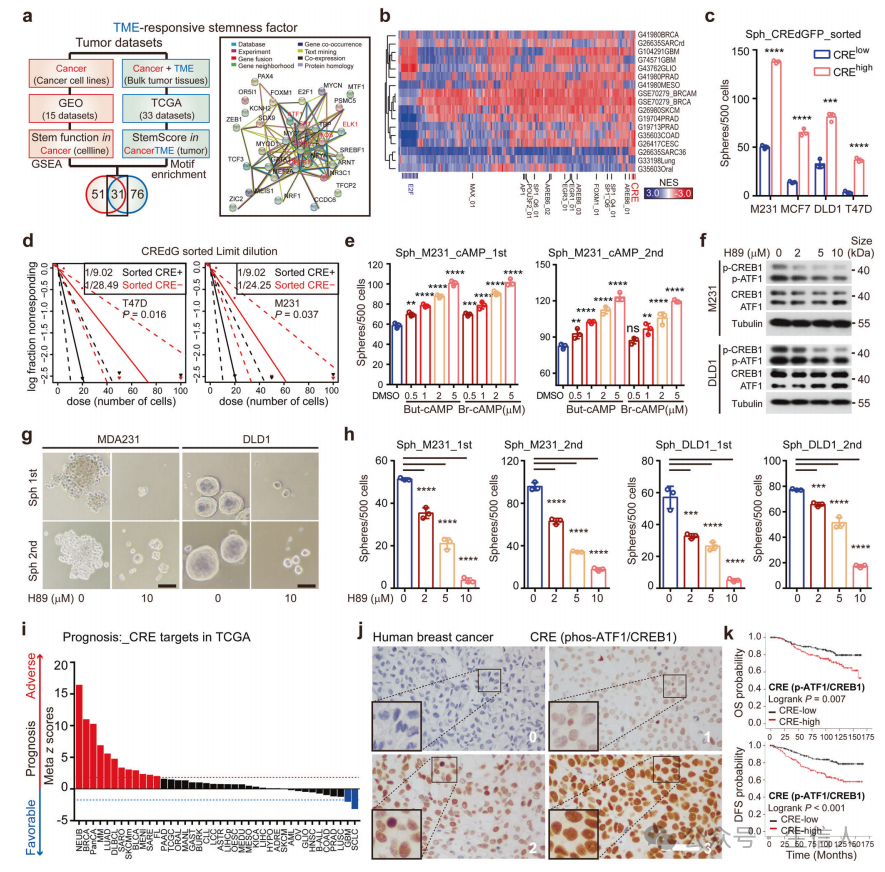
作者对33种TCGA癌症类型的10852个完整肿瘤的转录谱进行了表征,以确定在stemness评分高的肿瘤中控制高表达基因的转录因子为107个TME相关的stemness因子。
为了确定在功能上参与癌症干性的TME干性因子,重新分析了来自肿瘤圈研究的CSC调节因子, 发现CRE是一个保守的TME反应性癌症干细胞程序。
研究构建了一个转录报告基因,该转录报告基因由一个合成的CRE启动子(CRE-dGFP)驱动的不稳定GFP (dGFP),它在cAMP刺激剂(Butyl-cAMP和Forskolin)或抑制剂(SQ22536, Supplementary Fig. S2a)下如实报告CRE活性,通过研究发现 CRE活性是癌症发生所必需的。
接着,研究了 CRE活性的临床意义,发现CRE活性提示肿瘤发生、复发和临床预后不良。
2.TME肾上腺素能信号指导CRE依赖性肿瘤的发生
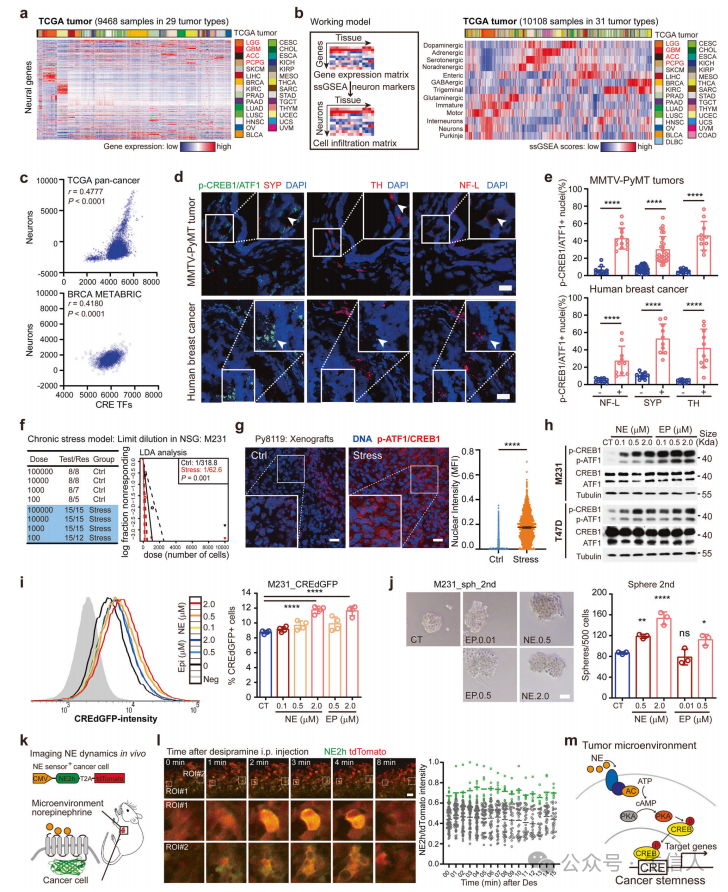
在细胞外分子中,神经信号是CRE活性的关键调节剂,与Stemness Scores呈正相关。同样,本研究也发现 CRE活动和干性功能都与TME神经信号有关。
为了确定TME神经信号与癌细胞CRE活性的空间关联,检测了神经标记物的定位和原位CRE活性, 发现TME肾上腺素能信号可能激活癌细胞中的CRE因子。接下来,确定了激活癌细胞中CRE转录因子的TME神经递质— 微环境局部化的去甲肾上腺素在激活CRE依赖的癌症干细胞中起关键作用。
确定了去甲肾上腺素在局部TME中的活性动态,发现了一种新的癌症干性模型,癌细胞利用TME神经信号获得CRE驱动的干性。
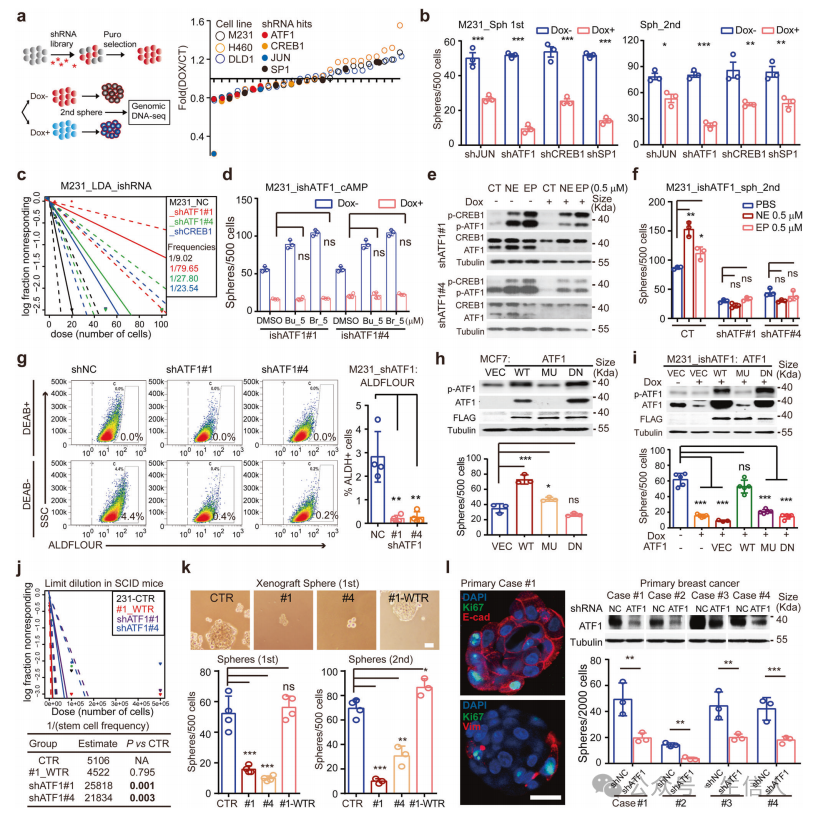
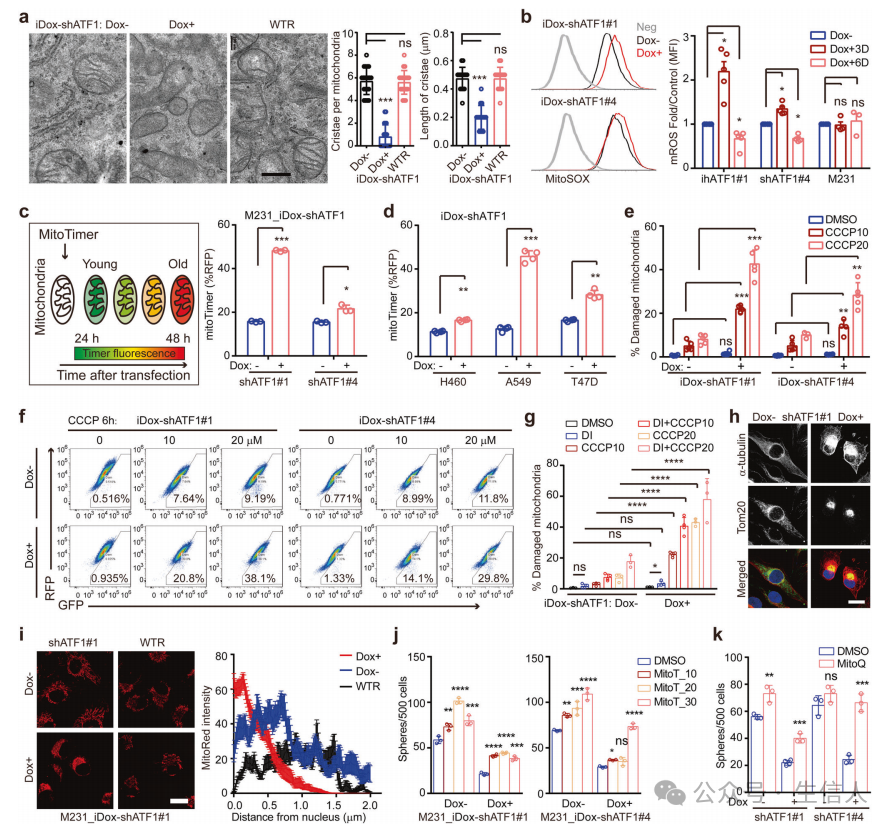
为了确定茎样功能所需的特定CRE因子,作者使用Dox诱导shRNA (iDox-shRNA)文库对序列球体形成进行了功能筛选,揭示了ATF1在神经信号cAMP介导的癌症发生中的重要作用。接下来评估了Dox+和Dox-washout环境下iDox-shATF1细胞的干样功能,ATF1在增强癌症干细胞中具有保守的、依赖于反式激活的作用。最后确认了ATF1在体内的功能, ATF1是一种特定的CRE转录因子,它指示TME神经信号,使癌症发生。
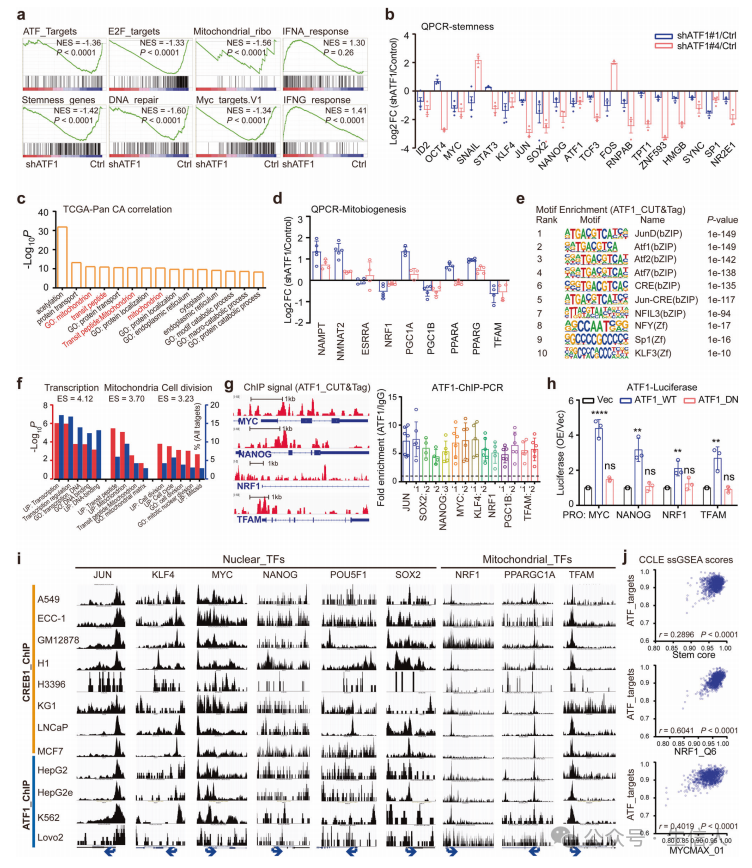
对iDox-shATF1 MDA-MB-231细胞进行了RNA测序,qPCR分析,对泛癌TCGA数据集中ATF1共表达基因的分析显示, 核多能性因子和线粒体生物发生调节因子都是ATF1的转录靶点。
为了确定ATF1与候选转录靶标的直接结合,进行了靶下切割和标记(CUT&Tag)测定,对ATF1免疫沉淀的染色质DNA进行PCR分析, 证实ATF1直接激活核多能因子和线粒体生物发生调节因子。
揭示了ATF1在反式激活多个癌细胞的核多能性和线粒体生物发生中的保守作用。转录组学分析表明,ATF1耗竭导致干细胞样癌细胞的衰老样表型。
为了确定ATF1如何控制线粒体状态,使用MitoTracker Red分析了iDoxshATF1细胞的线粒体活性,发现ATF1耗尽后线粒体慢性受损,,ATF1耗竭减少线粒体再生,诱导线粒体功能失调。 线粒体生物发生缺陷诱导线粒体功能失调,从而损害ATF1缺失细胞的癌症干细胞性。
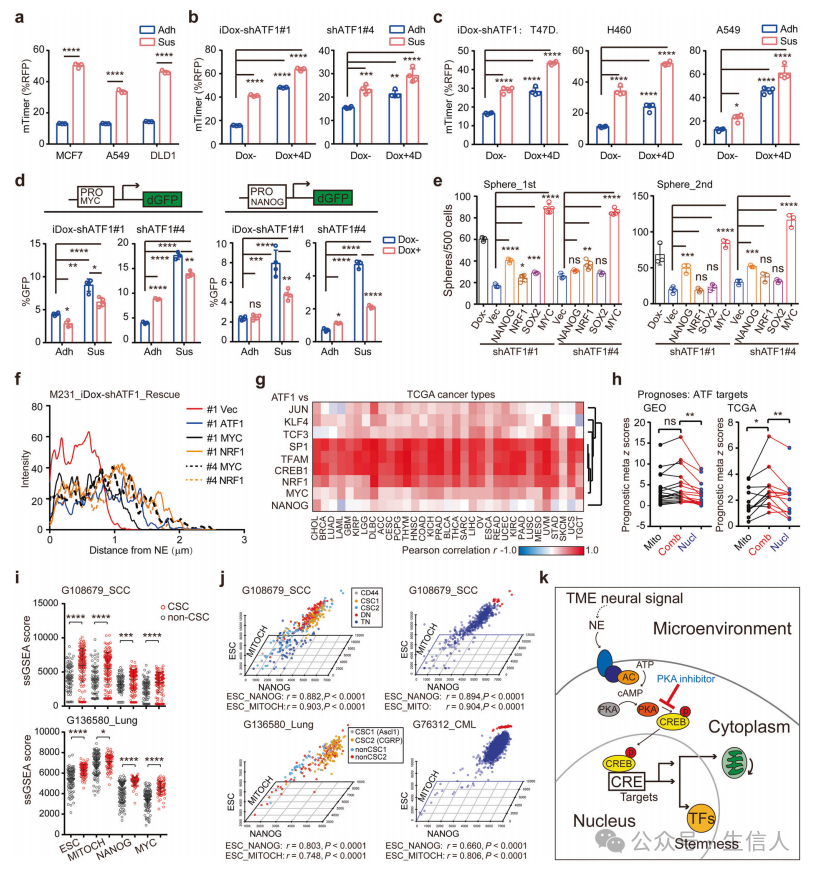
ATF1缺陷细胞的线粒体功能障碍表明,细胞适应性可能是ATF1定向干细胞计划的关键机制。使用试验验证这一假设,并且发现ATF1引导的核多能性和线粒体生物发生使对悬浮条件的适应性反应获得了干样性质。接下来,确定了核多能性和线粒体生物发生如何促进ATF1介导的癌症干细胞, 核多能性和线粒体生物发生的协同作用是ATF1依赖性癌症发生的基础。
为了确定ATF1在协调癌症中核多能性和线粒体调节因子中的临床相关性,评估了TCGA数据集中ATF1与候选基因之间的相关性。 个体癌症队列的生存分析证实了ATF1/CREB1共同表达核和线粒体靶点的患者预后较差,证实了ATF1协调核-线粒体程序的临床相关性。
为了破译单个CSCs中协调的核多能性和线粒体生物发生,使用单细胞转录组数据集评估了单细胞核多能性和线粒体生物发生水平, 由TME神经信号- ATF1通路引导的核多能性和线粒体生物发生的耦合激活是癌症干细胞样状态的保守机制。
图6:ATF1协调的核-线粒体状态是癌症发生的基础
本研究揭示了癌症细胞如何利用微环境中的神经信号来激活核与线粒体之间的协调,从而获得干细胞特性。通过大规模的泛癌症转录组筛选和一系列实验验证,作者发现 CRE转录因子在这一过程中扮演了关键角色。这一发现不仅有助于我们更深入地理解癌症的发生和发展机制,还为未来的癌症治疗提供了新的思路和方法。例如,通过抑制CRE转录因子的活性或阻断其与下游基因的相互作用,可能能够有效地抑制癌症细胞的干细胞特性和耐药性,从而提高癌症治疗的效果。
版权声明:本文为“乐问号”作者或机构在乐问医学上传并发布,仅代表该作者或机构观点,不代表乐问医学的观点或立场,不能作为个体诊疗依据,如有不适,请结合自身情况寻求医生的针对性治疗。
链接:http://www.lewenyixue.com/2024/07/26/%E9%AB%98%E5%88%86%E6%80%9D%E8%B7%AF%E4%B9%8B%E8%BF%9B%E9%98%B6%E7%89%88%E8%82%BF%E7%98%A4%E5%B9%B2%E6%80%A7%E5%8F%8C%E7%83%AD%E7%82%B9%E8%81%94%E5%90%88%E5%88%86%E6%9E%90/
THE END