Nat Immunol | 细胞形状响应引导树突状细胞向淋巴结迁移
免疫细胞在外周组织和淋巴器官之间的循环运动对于免疫应答的成功非常关键。在迁移过程中,免疫细胞骨架需要动态重组,响应外部信号 【1】 。趋化因子,尤其是CCR7,对维持免疫细胞在稳态、感染和癌症中的平衡至关重要 【2】 。生化信号、炎症介质和机械影响都可能改变CCR7的表达,从而影响免疫细胞的迁移。研究表明,机械破坏细胞间连接可以诱导树突状细胞 (DC) 中CCR7的表达 【3】 。然而,目前尚不清楚DC是否在外周组织中形成这种连接,并影响DC向淋巴结的迁移。
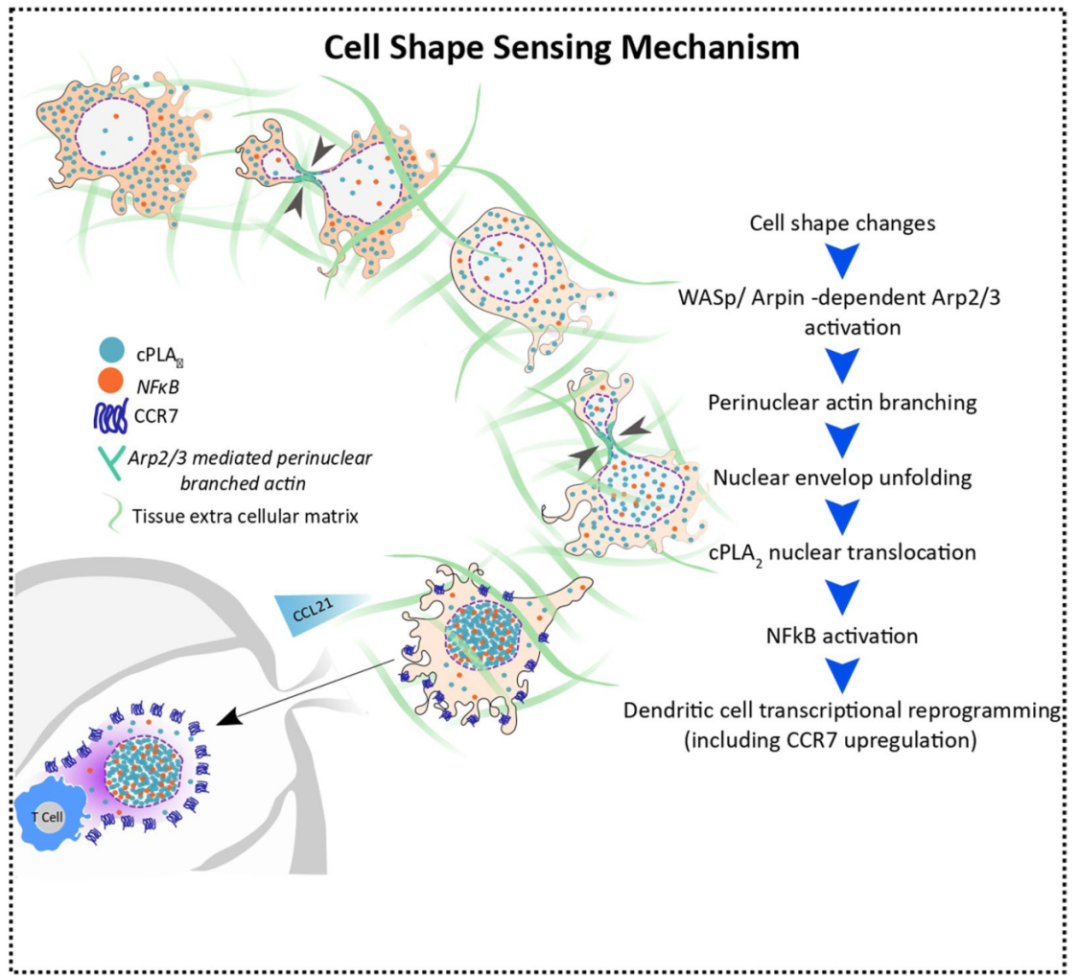
近日,来自法国巴黎文理研究大学居里研究所的 Ana-Maria Lennon-Duménil 与 Matthieu Piel 共同在 Nature Immunology 期刊发表题为 Cell shape sensing licenses dendritic cells for homeostatic migration to lymph nodes (细胞形状感知引导稳态条件下树突状细胞向淋巴结的迁移) 的文章。 研究揭示了免疫细胞通过形状感知机制调控趋化因子受体CCR7的表达,从而引导DCs从外周组织迁移到淋巴结。这一过程受到cPLA2-ARP2/3信号轴和IKKβ-NF-κB通路的影响。研究结果表明,免疫细胞的形态改变对其迁移行为和免疫调节功能产生影响,突显了组织物理特性在适应性免疫中的重要作用。
在组织中,受物理约束的运动细胞会发生形态变化和内部细胞器的变形,而免疫细胞能够适应并响应这种形态变化 【4】 。这些变化会引发核变形事件,激活脂质代谢酶细胞质磷酸脂酶2 (cPLA2) ,产生花生四烯酸 (AA) 并增强肌动蛋白的收缩能力,帮助细胞克服在密集组织中的阻力,使它们保持移动 【5】 。然而,目前尚不清楚cPLA2激活对免疫细胞的转录活动有何影响。
DC向淋巴结的迁移需要CCR7的表达来增强其运动能力。因此,研究人员使用细胞限制装置控制DC的形态变化,触发CCR7表达,促进DC运动。实验结果显示,将未成熟DC限制在3µm形变装置中可以显著增加CCR7的表达,促使细胞展现出趋化性和增强的运动能力,而2µm或4µm的形变装置则没有这种效果。研究人员观察到3µm形变装置可同时诱导DCs的CCR7表达和运动性,表明这两个过程可能受到共同机制的控制。进一步研究显示,在3µm形变装置中,抑制cPLA2酶活性能够阻止DC 中CCR7的上调,并观察到cPLA2的核内积累。这些研究结果提示,DC运动和CCR7表达的协同上调依赖于cPLA2的活性和核内积累,而且需要保持核膜的完整性。更重要的是,DCs表现出对细胞形态变化极为精确的感知机制。接下来,研究人员探究了导致cPLA2核转位和活化的机制。结果显示,在受限并经历形态变化的DC中,ARP2/3复合物在cPLA2核转位和活化中发挥关键作用。研究人员观察到,在3µm形变装置中,大约45%的细胞核周围存在肌动蛋白聚集,这一结构的维持依赖于ARP2/3活性。进一步研究发现,用ARP2/3抑制剂CK666处理DC或敲除ARP2/3激活蛋白WASp (WASp KO) 均可抑制ARP2/3的活性,调控DCs对形状变化的敏感性,并调节cPLA2的核积累阈值以及CCR7基因的表达。与之相反,敲除ARP2/3抑制因子Arpin则可增强ARP2/3的活性。这些结果揭示了DC对形状变化的敏感机制,为免疫细胞功能调节提供了重要线索。随后,研究人员通过探究核膜的折叠、张力以及支状肌动蛋白成核,阐述了细胞形态变化感知cPLA2 激活的分子机制。研究发现,ARP2/3 在控制核膜折叠和张力方面起着关键作用。实验结果表明,CK666 处理可以消除不同形变装置中DCs的核膜折叠差异。特别是在3µm形变装置的DCs 中,核膜周长显著减小,说明ARP2/3 抑制促进了核膜折叠。分析还表明,支状肌动蛋白成核有助于核膜的展开和张力调节,从而影响 cPLA2 的激活。核膜张力测量结果显示,CK666 处理可以降低核膜张力。缺乏层粘连蛋白的DCs无法在形变中产生足够的核膜张力,因此无法上调CCR7的表达,也无法完成cPLA2的核转位和激活。这些结果说明ARP2/3 在核膜展开和张力调节以及cPLA2的激活中发挥着重要作用。
细胞形状感应机制证明ARP2/3依赖性cPLA2的激活可能允许DC在稳态情况下迁移到淋巴结,而Arpin可能对这一过程起到负调节作用。动物实验显示,WASp KO和cPLA2KO小鼠中迁移至淋巴结的DC亚群cDC2s数量明显减少,而Arpin KO小鼠中的cDC2s数量明显增加。这些结果暗示,通过调节ARP2/3活性,WASp和Arpin可以控制cPLA2的激活阈值和CCR7的表达,从而精细地调节稳态条件下迁移至淋巴结的DCs数量。进一步研究发现,稳态下迁移至淋巴结的cDC2s表现出特定的转录谱,富含NF-κB和IFN相关基因,而IKKβ介导的NF-κB激活对于cDC2s的迁移是必需的。此外,3µm形变装置中,cPLA2 WT DCs许多基因被上调或下调,包括CCR7,而在cPLA2 KO DCs中,编码ARP2/3和IKKβ的基因以及Ccr7的表达上调完全消失,这说明CCR7的上调依赖于cPLA2。而且,IKKβ的抑制或缺失导致DCs中CCR7无法上调,但对cPLA2的核积累没有影响,这说明cPLA2作用于IKKβ和NF-κB的上游,诱导DC形状感知后的CCR7表达上调。这些发现揭示了依赖于IKKβ-NF-κB途径的ARP2/3-cPLA2细胞形状感应轴的激活推动了DCs迁移至淋巴结。研究还发现,DCs在稳态条件和炎症条件下向淋巴结的迁移存在显著差异,ARP2/3和cPLA2调控的CCR7上调对前者而言至关重要,但对后者并非必需。但是,无论是稳态条件还是炎症条件,DC迁移至淋巴结可能都需要IKKβ-NF-κB途径的参与。研究人员还观察到,受限DC中编码cPLA2和前列腺素E2 (PGE2) 受体的基因表达更加显著,这促使研究人员探究花生四烯酸 (AA) 产生的PGE2是否有助于受限DC的转录重编程。实验结果显示,PGE2能够部分弥补cPLA2的缺失,诱导NF-κB核转位。最后,研究人员比较了受限DC与LPS处理DC引发的信号通路。结果显示,在受限DC中“辅助T细胞分化调控”信号较弱,与T细胞应答有关的关键激活信号和细胞因子表达较少。此外,与LPS处理相比,受限DC在激活T淋巴细胞方面表现不足。分子机制研究揭示,受限DC激活了IRF1、STAT1、STAT3和STAT5A等转录因子,这种转录重编程使DC表现出特定的免疫调节状态。这些研究结果表明,细胞骨架和脂质代谢酶cPLA2之间的相互作用响应细胞形状改变,在转录水平重新编程DC,使其具备迁移至淋巴结的能力,同时赋予DC特定的免疫调节特性。
综上所述,该研究 揭示了DCs在特定形变条件下如何启动形态感知机制,通过ARP2/3-cPLA2-NF-κB途径调控CCR7的表达,从而影响其向淋巴结迁移,赋予其特定的免疫调节特性。研究表明DCs能够利用细胞骨架和脂质代谢的相互作用来适应环境中的物理约束。
https://doi.org/10.1038/s41590-024-01856-3
制版人:十一
1. Parsons, J. T., Horwitz, A. R. & Schwartz, M. A. Cell adhesion: integrating cytoskeletal dynamics and cellular tension. Nat. Rev. Mol. Cell Biol. 11, 633-643 (2010).
2. Förster, R., Davalos-Misslitz, A. C. & Rot, A. CCR7 and its ligands: balancing immunity and tolerance. Nat. Rev. Immunol. 8, 362-371 (2008).
3. Jiang, A. et al. Disruption of E-cadherin-mediated adhesion induces a functionally distinct pathway of dendritic cell maturation. Immunity 27, 610-624 (2007).
4. Raab, M. et al. ESCRT III repairs nuclear envelope ruptures during cell migration to limit DNA damage and cell death. Science 352, 359-362 (2016).
5. Venturini, V. et al. The nucleus measures shape changes for cellular proprioception to control dynamic cell behavior. Science 370, eaba2644 (2020).
版权声明:本文为“乐问号”作者或机构在乐问医学上传并发布,仅代表该作者或机构观点,不代表乐问医学的观点或立场,不能作为个体诊疗依据,如有不适,请结合自身情况寻求医生的针对性治疗。
链接:http://www.lewenyixue.com/2024/07/02/Nat%20Immunol%20%7C%20%E7%BB%86%E8%83%9E%E5%BD%A2%E7%8A%B6%E5%93%8D%E5%BA%94/
THE END






赶快来坐沙发