



Nat Immu | 脓毒症训练的巨噬细胞促进抗肿瘤组织驻留T细胞
撰文:拾人间烟火
IF=30.5
推荐度: 
亮点:
败血症这种失调的宿主反应属于全身炎症反应综合征( SIRS ),其特征是过度释放炎症细胞因子,激活先天和获得性免疫。因此,败血症可诱导免疫系统发生深刻的改变长达数月,在这期间,要么免疫耐受性增强要么先天免疫得到驯化。在此,作者假设,在脓毒症幸存者中观察到的免疫重编程可以改变患癌症的风险。首先,作者在真实世界的医疗记录中检验了这一假设,这些记录来自法国国家行政部门关于住院的健康索赔数据。通过这些数据,研究发现败血症幸存者的癌症累计发病率低于相应的非严重感染幸存者。
2024 年 4 月 29 日,法国南特大学 Antoine Roquilly 团队在 Nature Immunology 上发表题为 “Sepsis-trained macrophages promote antitumoraltissue-resident T cells” 的文章。

败血症引起免疫改变,这种改变在疾病消退后持续数月。这种免疫重编程对患癌症风险的影响尚不清楚。在这里,使用一个国家索赔数据库来显示败血症幸存者的癌症累积发病率低于匹配的非严重感染幸存者。确定了脓毒症训练的巨噬细胞释放的趋化因子网络,该网络通过 CCR2 和 CXCR6 刺激触发 T 细胞的组织驻留,作为脓毒症治愈后降低新发肿瘤风险的免疫机制。虽然非感染性炎症不会引起这一网络,但层粘连蛋白注射可以治疗性地再现保护性脓毒症的作用。在脓毒症患者中检测到这种趋化因子网络和 CXCR6 组织驻留 T 细胞积累,并与癌症患者的延长生存期有关。这些发现确定了与治疗相关的败血症诱导的训练免疫的抗肿瘤结果。
感染是全世界最常见的死亡原因之一。败血症被定义为由宿主对感染反应失调引起的危及生命的器官功能障碍,是大多数感染相关死亡的原因。这种失调的宿主反应属于系统性炎症反应综合征 (SIRS) ,其特征是过度释放炎症细胞因子,激活先天和获得性免疫。值得注意的是, SIRS 也可以由非无菌条件引起,如严重创伤或大手术。
随着医疗管理水平的提高,败血症相关死亡的风险在过去几十年里稳步下降。因此,脓毒症后存在潜在长期后遗症的患者数量增加,心血管疾病和痴呆的风险增加。
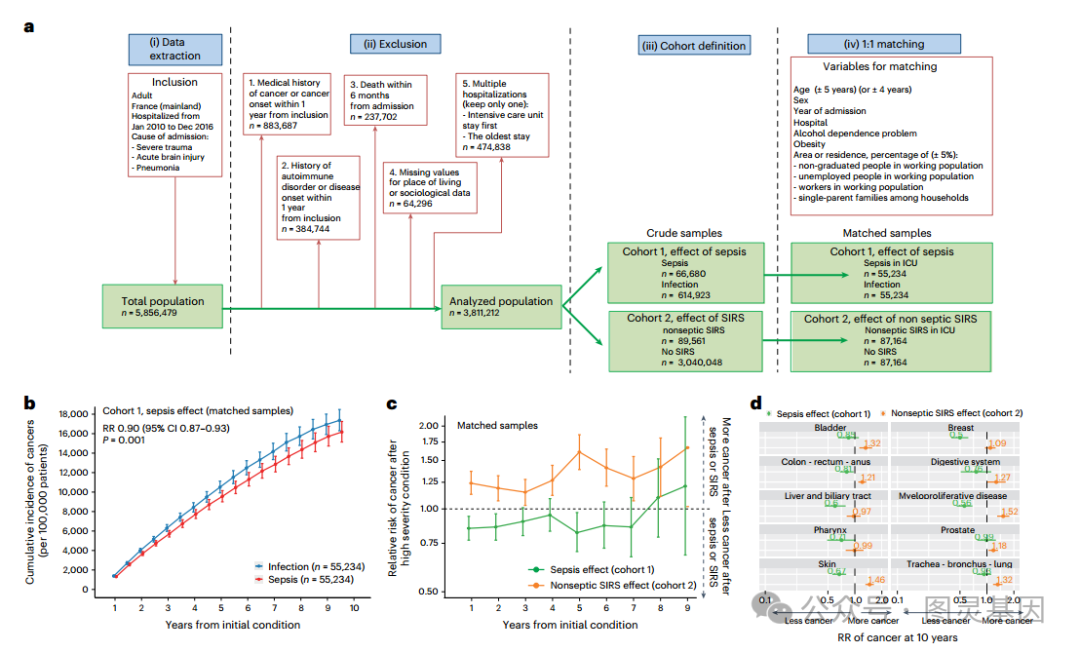
图 1 人类败血症治愈后 10 年内癌症风险降低
脓毒症会诱发长达数月的免疫系统改变,要么增加免疫耐受性,要么训练先天免疫 ; 然而,长期免疫重编程对脓毒症治愈患者发生新发癌症的长期风险的影响尚未得到彻底研究。研究人员假设,在败血症幸存者中观察到的免疫重编程可以改变患癌症的风险,质疑败血症和非败血症 SIRS 对长期预后是否有相同的影响。首先,在现实世界的医疗记录中检验了这一假设,这些记录来自法国国家行政卫生索赔住院数据。因此,研究败血症后的免疫重编程可能揭示免疫耐受或癌症控制的机制特征。研究结果全面描述了败血症对免疫的长期影响,并揭示了驻留巨噬细胞调节 T 细胞驻留的能力, T 细胞作为肿瘤免疫监视的关键介质。
1. 降低败血症幸存者患癌症的风险
为了研究脓毒症诱导的人类长期免疫改变的影响,比较了脓毒症存活患者与相关匹配对照患者发生新癌症的风险。为此,使用法国国民保险和行政数据库系统来比较 2010 年 1 月至 2016 年 12 月住院患者中败血症时不存在的癌症的发展情况,并于 2020 年 12 月结束随访。法国医院的活动被全面纳入法国医院出院数据库 (PMSI) ,作为全国所有住院患者的完整记录。
法国共有 5,856,479 名成年人因严重创伤、急性脑损伤或肺炎住院 ( 图 1a) 。排除了有癌症病史 (n = 883,687) 或自身免疫性疾病 (n = 384,744) 、在指标入院后 6 个月内死亡的成年人,因为这可能与未治愈的初始状况 (n = 237,702) 或缺少匹配所需的社会学数据 (n = 64,296) 有关。对于研究期间多次住院的患者,保留了最早的重症监护住院,如果适用,或者保留了最早的住院 (n = 474,838 次住院被删除 ) 。
最终定义了两组来研究脓毒症 ( 队列 1,n = 681,603) 和非脓毒症 SIRS( 队列 2,n = 3,219,609) 对癌症风险的影响。在队列 1 中纳入了脓毒症 ( 社区或任何医院获得性感染,因危及生命的情况住院重症监护病房 ) 或感染 ( 社区或任何医院获得性感染,住院普通病房,无急性器官衰竭 ) 的患者 ; 队列 2 中的非感染性 SIRS 患者 ( 因严重创伤或脑损伤住院,无感染,因危及生命的情况住在重症监护病房 ) 和非 SIRS 患者 ( 在普通病房住院,住院期间无感染或急性器官衰竭 )( 图 1a) 。因此,能够研究脓毒症与感染幸存者 ( 脓毒症效应 ) 以及非脓毒性 SIRS 与非 SIRS 幸存者 ( 非脓毒性 SIRS 效应 ) 在出院后 10 年内对癌症风险的影响 ( 总风险和原因 ) 。
出院后 10 年,脓毒症幸存者与感染幸存者相比,新癌症发展的累积风险更低 ( 相对风险 (RR) 0.93, 95% 可信区间 (CI) 0.91-0.95; 图 1a) 。为了大大减少与观察性研究相关的偏倚,考虑了患者人口统计学 ( 年龄、性别、肥胖、酗酒和入院原因 ) 、社会 ( 居住地编码 ) 和环境 ( 医院编码定位 ) 癌症危险因素在组间的差异,以 1:1 的比例进行败血症幸存者与感染治愈患者的个体匹配。在匹配的比较中,败血症幸存者的累积癌症发病率仍然低于感染幸存者 (RR 0.90, 95% CI 0.87-0.93; 图 1 b) 。更好地考虑了混杂因素的残余影响,用 Cox 模型计算出的已知危险因素 ( 匹配标准 ) 校正后的癌症风险比 (HR) 为 0.93 (95% CI 0.88-0.98) 。
在敏感性分析中,采用了严格的匹配标准, HR 为 0.87 (95% CI 0.83-0.91; 图 1b) 。在亚组分析中,严重程度评分较高的脓毒症患者 ( 简化严重程度指数 2 (IGS-II) > 25;RR 0.77;95% CI 0.70-0.85) ,但在低严重性脓毒症患者中没有 (IGS-II 15-25, RR 0.98;95% CI0.94-1.02) ,表明脓毒症严重程度与癌症风险降低之间存在剂量效应关系。当纳入随访前 6 个月死亡的患者 ( 调整后危险度 0.91,95% CI 0.88-0.94) 或随访期间诊断出自身免疫性疾病的患者 ( 调整后危险度 0.94,95% CI 0.91 - 0.97) 时,败血症后癌症风险的估计几乎没有改变。根据癌症的瞬时 RR 估计,脓毒症对癌症发生风险的影响在前 7 年的随访中是恒定的,然后逐渐降低 ( 图 1c 在匹配人群样本中,图 1d 在粗人群中 ) 。
通过估计 10 个最常见癌症部位的 RRs ,脓毒症的影响不是癌症特异性的 ( 图 1d) 。值得注意的是,与烟草无关的癌症,如结肠癌 - 直肠 - 肛门癌、肝癌、皮肤癌和乳腺癌,都在这一名单之列,这反驳了烟草使用不平衡的偏见。
质疑 SIRS 的保护作用是否仅限于脓毒症,或者是否可以在非脓毒症 SIRS 后观察到,如队列 2 。与败血症相反,非败血性 SIRS 与随访 10 年后癌症发病率增加相关。这种效应在整个研究期间保持不变 ( 图 1c) ,并且不是癌症特异性的 ( 图 1d) 。这些数据强化了败血症后终生免疫重编程的假设,并提示败血症和非败血症 SIRS 之间的差异。
2. 败血症治愈小鼠控制肺癌的数量、大小和生长
尽管匹配策略限制了癌症不平衡危险因素的偏倚风险,但其他未控制的混杂因素仍可能是脓毒症后观察到的癌症风险降低的原因。因此,目的是在对照小鼠模型中测试脓毒症治疗对癌症易感性的影响。肺癌是全世界癌症死亡的主要原因,而下呼吸道感染是脓毒症最常见的原因,这表明研究肺脓毒症后肺癌的生长具有医学意义。使用了非致死性大肠杆菌肺炎模型,该模型诱导急性低氧血症性呼吸衰竭、乳酸酸中毒和与先天免疫激活相关的体重减轻,因此符合败血症的定义。
在这个非致死性脓毒症模型中,小鼠自发清除了细菌,在第 7 天观察到体重和组织结构正常化。因此,小鼠从第 7 天就被认为治愈了脓毒症,但在脓毒症后 28 天注射时,同基因 Lewis 肺癌 (LLC) 的发展减少,并在脓毒症后 56 天逐渐恢复正常 ( 图 2a) 。更准确地说,在脓毒症治愈的动物中观察到肿瘤的绝对数量和肿瘤大小的减少。质疑败血症治愈小鼠肿瘤生长的减少是否仅限于肺肿瘤,是否可以在其他组织中观察到。因此,在预先验证的小鼠败血症自发治愈模型中测试了黑色素瘤 B16-OVA( 一种产生同基因皮肤癌的卵清蛋白 (OVA)) 的原位皮肤生长情况。观察到败血症治愈小鼠的皮肤黑色素瘤肿瘤生长减少 ( 图 2b) ,表明这种现象并不局限于肺部。
LLC 和黑色素瘤 B16-OVA 是免疫原性的,这意味着当癌细胞分别注射到肺部和皮肤时,免疫细胞会减少肿瘤的生长。质疑当癌症免疫原性差时,败血症是否仍能抑制肿瘤生长。为了解决这个问题,研究了由 ova 阳性 (OVApos) 大肠杆菌或 ova 阴性 (OVAnull) 大肠杆菌引起的肺脓毒症治愈的小鼠中黑色素瘤 B16-OVA 的肺转移发展,其免疫原性在这种异位定位中下降 13( 图 2c) 。在该模型中, T 细胞被 OVApos 大肠杆菌引物,识别 OVApos 肿瘤细胞,从而恢复免疫原性。在 OVAposE 治愈的小鼠中,肺黑色素瘤转移的发展减少。但在单独用 OVA 治疗或 OVAnullE 治愈的小鼠中没有。大肠杆菌 ( 图 2c) 。这一系列实验表明,脓毒症治愈小鼠模型再现了在人类中观察到的癌症生长下降,并表明这种现象可能是免疫介导的。

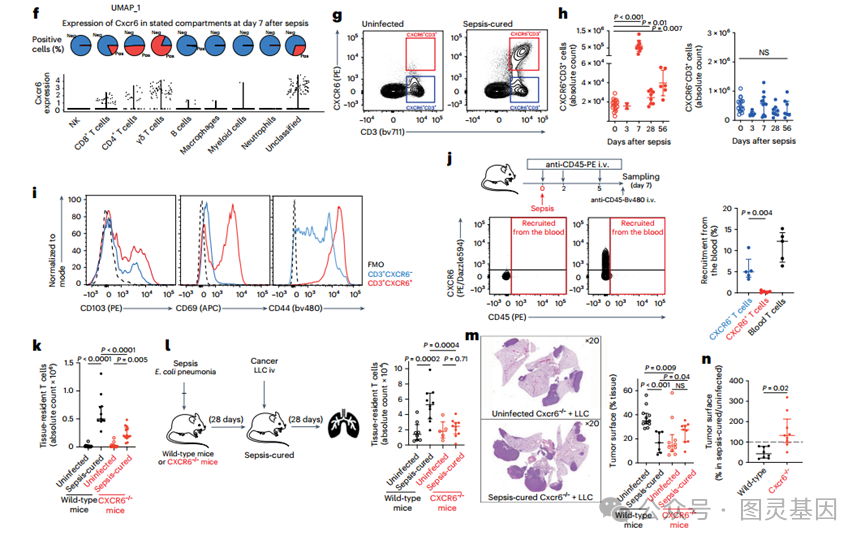
图 2 CXCR6+ 组织驻留 T 细胞减少败血症治愈小鼠的癌症生长
3. 脓毒症后 CXCR6+ 组织驻留 T 细胞的局部积累
然后,分析了整个肺脓毒症的单细胞免疫景观,以确定脓毒症后减少癌症发展的分子和细胞改变。为此,采集了未感染小鼠以及脓毒症后 1 、 3 和 7 天的肺免疫细胞,分别代表未感染状态 ( 第 0 天 ) 、脓毒症急性期 ( 第 1 - 3 天 ) 和脓毒症治愈状态 ( 第 7 天 ) 。分析包括 10,542 个细胞,经过质量控制,标签解复用和去除双重,从两个独立的实验中取样,每个时间点有两只小鼠。基于差异表达最多的基因手动注释细胞簇 ( 图 2d) 。早在第 1 天就观察到免疫细胞群的快速重组。大肠杆菌肺炎期间 ( 第 1 天和第 3 天 ) 的免疫改变包括中性粒细胞频率的显著增加,这与败血症期间的预期一致 ( 图 2e) 。
中性粒细胞频率在第 7 天恢复到基线值,证实败血症消退。在败血症治愈状态下 ( 第 7 天 ) ,在 T 细胞中观察到最明显的免疫室变化,特别是 Cxcr6+ T 细胞 ( 表达 Cxcr6 、 CD3g 和 Il17 的前 5 个基因 ) 和巨噬细胞表达 Cxcl16 (Cxcr6 配体 ) 的出现 ( 图 2e) 。 Cxcr6 的表达仅限于 T 细胞 ( 图 2f) ,通过流式细胞术证实,在败血症治愈小鼠的肺中, Cxcr6 + T 细胞数量的增加持续了 56 天 ( 图 2g,h) 。值得注意的是, CXCR6 − T 细胞的数量变化不大,表明败血症后 T 细胞的积累依赖于 CXCR6 。
4. 脓毒症后肿瘤发展的 cxcr6 依赖性降低
CXCR6 是组织常驻 T 细胞的标记物,以其介导细胞毒性和细胞因子反应以及增强癌症免疫控制的能力而闻名。研究人员证实,脓毒症治愈小鼠积累的 CXCR6+ T 细胞具有 CD103 (Itgae) 、 CD69 和 CD44 的高膜表达 ( 图 2i) ,代表了组织驻留 T 细胞的典型特征 17 。 Cxcr6+ T 细胞具有高组织驻留 T 细胞一致转录组特征,特别是低 Ccr7 和 S1pr1 表达水平。为了验证 CXCR6+ T 细胞来源于组织前体,通过静脉注射 (i.v.) 植红蛋白 (PE) 偶联抗 cd45 标记体内循环免疫细胞。研究人员观察到,在脓毒症期间,约 15% 的 CXCR6 − T 细胞从血液中被募集到肺部,但 CXCR6+ T 细胞仍然是 PE - 证实这些细胞不是通过血液募集,而是来自局部祖细胞 ( 图 2j) 。
因此,假设败血症后观察到的肿瘤生长减少是由组织驻留 T 细胞的 cxcr6 依赖性积累介导的。为了支持 CXCR6 的作用,观察到 CXCR6 缺陷小鼠败血症后组织驻留 T 细胞的数量低于野生型小鼠 ( 图 2k) 。研究人员还注意到,脓毒症在 LLC 发育过程中增加了组织驻留 T 细胞的数量,但在 cxcr6 缺陷小鼠中没有 ( 图 21) 。 cxcr6 缺陷小鼠脓毒症后 LLC 表面没有减少,与野生型脓毒症治愈小鼠相比, LLC 表面增加 ( 图 2m) 。这一系列实验表明,脓毒症对野生型和 cxcr6 缺陷小鼠的肿瘤生长具有相反的影响 ( 图 2n) 。由于 CXCR6 的表达仅限于组织常驻 T 细胞,这一观察结果提示 CXCR6+ 组织常驻 T 细胞在这一现象中的作用。
5. 脓毒症减少增殖的 CXCR6+ T 细胞的组织出口
然后,研究了脓毒症后观察到的 CXCR6 组织驻留 T 细胞增加的原因。发现增殖标志物 Ki67 在 CXCR6+ 中上调,而在 CXCR6 - 常驻 T 细胞中没有上调 ( 图 3a) 。组织出口抑制也是诱导增殖 T 细胞驻留的关键机制。为了测试败血症是否会增加肺组织诱导 T 细胞驻留的能力,将从未感染的 CD45.2pos 野生型小鼠收集的 T 细胞转移到未感染或败血症治愈的 CD45.1pos 受体的肺部 ; 然后,在 36 小时后测量受体肺中转移的 T 细胞的数量。
研究人员观察到,在败血症治愈的受体小鼠中,来自转移细胞的常驻 T 细胞保留在肺部的数量增加 ( 图 3b) 。从这一系列的实验中,研究人员得出结论,败血症留下了免疫组织重组,这减少了局部增殖 T 细胞的组织出口。
研究人员进一步表征了这些 CXCR6+ 组织驻留 T 细胞,并观察到在未感染小鼠中约 50% 是 CD8+ ,而在脓毒症治愈小鼠中,大多数属于 γδ T 细胞和 CD4+ T 细胞 ( 图 3c,d) 。重要的是,这一观察结果与这些细胞亚群上 CXCR6 的表达水平相关,而在败血症治愈小鼠中, γδ 、 CD4+ 和 CD8+ CXCR6 − 组织常驻 T 细胞的数量保持不变。研究人员观察到,在未感染小鼠中, CXCR6+ γδ T 细胞平均分布在 v - γ1 、 v - γ4 和 v - γ6 链 (Heilig 和 Tonegawa 命名法 ) 之间,而在败血症治愈小鼠中, CXCR6+ γδ T 细胞主要是 IL-17+ r - γ T + v - γ6δ T 细胞 ( 图 3e,f) 。还通过单细胞 TCR 测序 (scTCR-seq) 研究了败血症对 TCR-β 库的影响,并观察到在败血症治愈小鼠的 CXCR6+ T 细胞中发现了最常见的克隆型 ( 图 3g,h) 。
研究人员得出结论,脓毒症诱导 CXCR6+ 原位寡克隆扩增然后质疑这种 TCR 选择是否调节了它们的功能。为了解决这个问题,比较了克隆选择 ( 至少四个细胞表达最常见的克隆型 ) 和非克隆选择的 CXCR6+ 组织驻留 T 细胞 ( 仅在一个细胞中观察到克隆型 ) 细胞的转录组学特征 ( 图 3i) 。该方法发现,在克隆选择和非选择的 T 细胞之间,只有 9 个差异表达基因 (deg) ,其中 6 个与 TCR 直接相关,这表明对病原体的直接识别不会诱导其转录活性的持续改变。还研究了败血症对克隆选择 T 细胞产生 IFNγ 的影响。
为此,研究人员通过气管内注射卵清蛋白 (i.t) 和 CD45.2pos ova 特异性 OT-I 细胞 (i.v) ,诱导 CD45.1pos 小鼠形成 ova 特异性 CD8+ 组织驻留记忆 T 细胞。在诱导 OVA 特异性组织驻留 CD8+ T 细胞两周后,在小鼠中诱导大肠杆菌肺炎, 7 天后,在体内分析了对 i.t OVA 攻击的免疫反应 ( 图 3j) 。
研究人员观察到败血症增加了先前存在的 OVA 特异性 CD8+ 组织驻留 T 细胞的数量,但没有改变 IFNγ 的产生,以响应 OVA 刺激 ( 图 3j) 。研究人员得出结论,脓毒症治疗通过增殖 T 细胞的局部保留增加了寡克隆 CXCR6+ 组织驻留 T 细胞 ; 然而,败血症并没有增加他们在再次感染时对抗原的反应。
6. 脓毒症后 CXCR6 和 ccr2 依赖性 T 细胞组织潴留
因此,研究人员解读了 tcr 非依赖性肺微环境介质在败血症后积累组织驻留 CXCR6+ T 细胞中的作用。从单细胞转录组学图谱中识别受体 (R) - 配体 (L) 对是揭示塑造细胞命运的中心信号机制的有力工具。在 scRNA-seq 分析中选择了 Cxcr6+ T 细胞簇中基因表达最高的 15 个受体 ( 图 2e) ,然后进行基于文献的研究,评估其配体是否在任何免疫细胞中表达 ( 图 4a) 。为了将这一文献分析扩展到可用数据集中所有潜在的 R - L 相互作用,研究人员用生物信息学方法过滤了 Cxcr6+ T 细胞群中表达的所有 R ,并将 L 与其在所有细胞中的表达谱和脓毒症时间点相关联,以创建脓毒症期间和之后的 R - L 图谱。
研究人员根据它们的表达相关性选择了 L-R 对,从而得到了细胞间 L-R 通信的图形表示。在脓毒症期间 ( 第 3 天 ) ,这种无监督的方法定义了 Cxcr6-Cxcl16 对 (L 主要由巨噬细胞和树突状细胞表达 ) 和 IfnγR1-Ifnγ (L 主要由自然杀伤 (NK) 细胞表达 ) 对和 Ccr2-Ccl2 /Ccl7/ Ccl12 对 ( 来自 CellChat (KEGG mmu04060) 的 L 列表,均由巨噬细胞产生 ) 。脓毒症治愈后,又发现了 5 对,包括主要由巨噬细胞表达的 Tgf βRII-Tgfβ1 /Gdf15 (L) 树突状细胞 )( 图 4b) 。
研究人员接下来的目标是功能性地确认这些 L-R 对在败血症后建立 T 细胞组织驻留中的作用。为了优先探索 Cxcr6+ T 细胞特异性的 L-R 对,研究人员比较了 scRNA-seq 中定义的五个 T 细胞簇之间细胞内趋化因子信号传导基因的表达 ( 图 4c) 。研究人员观察到,在脓毒症的第三天, Cxcr6+ T 细胞簇中的 TGFβ 信号被激活,然后在第 7 天下降 ; 然而, TGFβ 信号通路在第 9 和 17 簇中也被激活,这表明 TGFβ 在败血症后对 CXCR6+ 组织驻留细胞没有特异性作用。当选择 ifn - γ r1 - IFNγ 对时,在 Cxcr6+ T 细胞簇中,干扰素 II 型信号通路在败血症期间和之后的激活程度低于其他 T 细胞簇,并且在 IFNγ 受体缺陷小鼠中, Cxcr6 组织内 T 细胞的数量没有减少。
因此,研究人员深入研究了 CXCR6 和 CCR2 受体,因为在 T 细胞簇中,它们的信号通路在 CXCR6 + 细胞中被选择性激活 ( 图 4c) ,它们在 T 细胞中的表达水平相关 ( 图 4d) 。首先,研究人员观察到,与野生型小鼠相比,败血症治愈的 Cxcr6 和 ccr2 缺陷小鼠的组织常驻 γδ T 和 CD4+ T 细胞数量减少 ( 图 4e,f) 。研究人员发现,在缺乏 cxcr6 的败血症治愈小鼠肺中的组织驻留 T 细胞中,增殖标记物 Ki67 的表达降低 ( 图 4) 。脓毒症后, cxcr6 缺陷小鼠纵隔淋巴结中组织常驻 T 细胞的数量高于野生型小鼠,这表明 cxcr6 缺陷的 T 细胞仍能在体内增殖,但不能在组织中保留 ( 图 4h) 。在缺乏 Ccr2 的败血症治愈小鼠中未见 Ki67 减少 ( 图 4i) 。
然后,研究人员的目标是在体内功能性地确认 T 细胞表达 CCR2 和 CXCR6 的作用。 CCR2 表达与先天免疫细胞共享。因此,将从未感染的 Cxcr6 或 Ccr2 缺陷供体小鼠中收集的 T 细胞转移到野生型受体中,以产生条件敲敲条件,其中只有转移的 T 细胞是 Ccr2 或 Cxcr6 缺陷的。与野生型 T 细胞转移所观察到的结果相反 ( 图 3b) ,脓毒症并没有增加肺中 CXCR6 缺陷或 CCR2 缺陷 T 细胞的数量 ( 图 4j) ,这表明脓毒症后 T 细胞表达 CXCR6 和 CCR2 对于其组织保留是必要的。此外,研究人员质疑 CXCR6 是否只需要启动 T 细胞的驻留或维持。为了回答这个问题,将从败血症治愈的野生型或 cxcr6 缺陷 CD45.2pos 小鼠肺部收集的常驻 T 细胞转移到未感染 CD45.1pos 受体的肺部。研究人员观察到缺乏 CXCR6 的转移细胞的保留减少 ( 图 4k) ,表明需要 CXCR6 来维持 T 细胞的组织驻留。
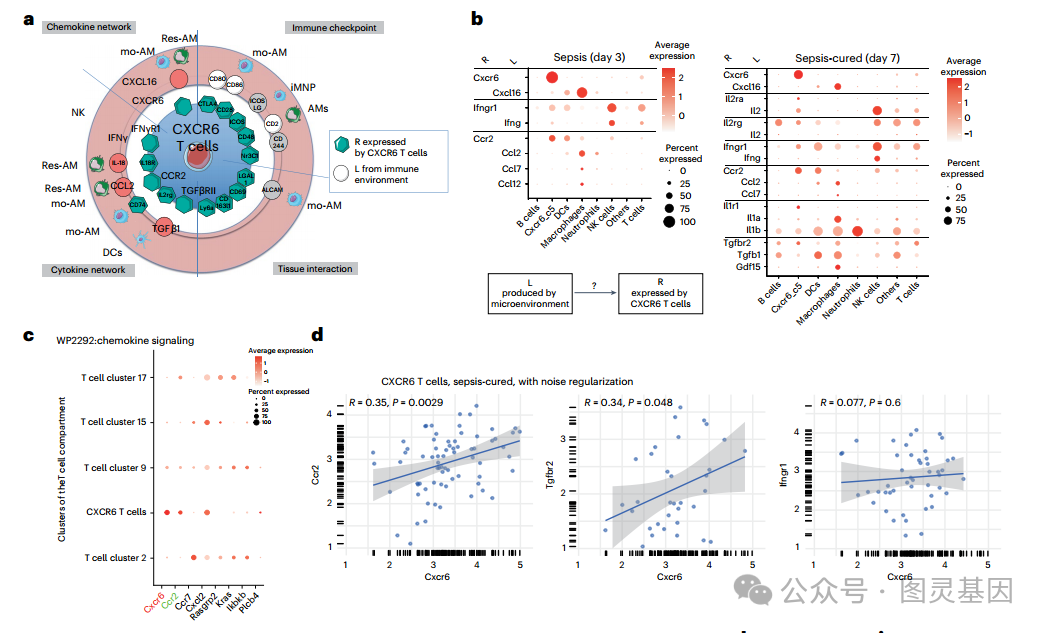
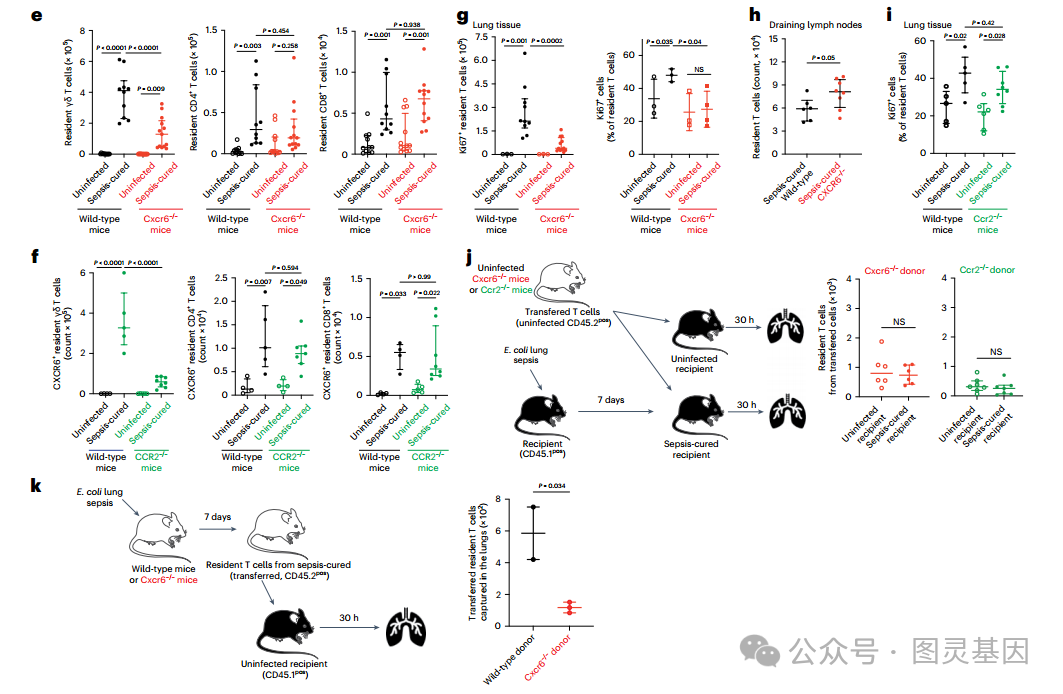
图 3 脓毒症治愈后增殖 T 细胞的 CXCR6 和 ccr2 依赖性组织保留
7.CXCR6 通过组织驻留 T 细胞调节细胞毒性和 Ccr2 的表达
通过对野生型和缺乏 CXCR6 的脓毒症治愈小鼠分选的常驻 T 细胞进行 scRNA-seq 分析,研究了 CXCR6 对常驻 T 细胞功能的影响。根据它们的转录谱定义了 γδ 、 CD4+ 和 CD8+ T 细胞,并根据它们的激活状态 (CD69 蛋白的低表达和高表达 ; 图 5a) 。 Cxcr6 缺乏主要影响 γδ T 和 CD4+ T 细胞百分比 ( 图 5b) 。 Cxcr6 缺乏显著影响了 γδ 常驻 T 细胞的转录活性,显著降低了 Ccr2 的表达水平 ( 图 5c) 。值得注意的是, Itgae (CD103) 和 Nrp1 (neuropilin) ,这两个 T 细胞驻留程序的成员,以及 Ctla2a( 细胞毒性 T 淋巴细胞相关蛋白 2α) 是 cxcr6 缺陷细胞中下调最多的基因。基因本体和基因集富集分析显示, Cxcr6 缺乏降低了驻留 T 细胞的迁移、趋化、细胞毒性和细胞杀伤功能 ( 图 5d) ,为 Cxcr6 + 组织驻留 T 细胞在脓毒症后抑制肿瘤生长的能力提供了合理的机械解释。
8.CCR2 增加 T 细胞中 CXCR6 的表达
Ccr2 是野生型和 Cxcr6 缺陷 T 细胞差异表达的主要基因之一 ( 图 5c) ,其在 γδ T 细胞中的表达水平与 Cxcr6 相关 ( 图 5e) 。研究人员还观察到, CCR2+ CXCR6+ 组织常驻 T 细胞 ( 双阳性 ) 的百分比在败血症期间增加 ( 图 5f,g) ,表明 T 细胞对其表达的共同调节。利用增强的绿色荧光蛋白 cDNA 取代 Cxcr6 编码区的 Cxcr6 缺陷小鼠,研究人员观察到,与未感染的受体相比,将 T 细胞转移到败血症治愈小鼠体内时, Cxcr6 报告基因的表达增加 ( 图 5h) 。研究人员还观察到,在 ccr2 缺陷小鼠中,常驻 T 细胞中的 CXCR6 表达水平降低 ( 图 5i) 。为了验证 CCR2 调节 CXCR6 表达的假设,研究人员将 CCR2 缺陷 T 细胞转移到败血症治愈野生型小鼠的肺中,观察到转移的 CCR2 缺陷 T 细胞中 CXCR6 蛋白水平降低 ( 图 5j); 然而,体外用 CCR2 激动剂处理幼稚 T 细胞并没有增加培养物中 CXCR6 的表达。因此,研究人员得出结论,通过 CCR2-CXCR6 通路的相互作用,脓毒后肺生态位使 T 细胞更适合居住,而共刺激因子可能参与了这一过程。
9. 增加 T 细胞驻留是由驻留 am 介导的
接下来,研究人员搜索了 CXCR6 和 CCR2 配体的细胞来源。为了支持 CXCL16 的作用, CXCL16 阻断抗体减少了组织驻留的 CXCR6+ T 细胞的总数,主要是 γδ T 细胞,但没有改变 CXCR6 − T 细胞 ( 图 6a) 。阻断 CXCL16 也降低了增殖标志物 Ki67 的表达。在使用 CXCL16 阻断抗体治疗的败血症治愈小鼠的 LLC 发展过程中,观察到 CXCR6+ 组织驻留 T 细胞的类似减少 ( 图 6b) 。
然后,研究人员质疑哪些细胞类型是败血症治愈小鼠中 CXCL16 的主要来源。研究人员观察到巨噬细胞是脓毒症治愈后 CXCL16 的唯一来源 ( 图 6c) ,也是组织常驻 T 细胞上表达的受体的主要配体来源。然后,检查了产生 cxcl16 的巨噬细胞是常住肺泡巨噬细胞 (Res-AMs) 还是脓毒症后重新填充气道的单核细胞源性巨噬细胞 (mo-AMs) 。验证了一种门控策略,使 Res-AMs 与 mo-AMs 能够分化,并发现大多数 AMs 在败血症后 28 天具有常驻表型 ( 图 6d) 。此外,观察到在第 7 天 CXCL16+ mo-AM 的数量增加,并迅速减少,在第 28 天, CXCL16+ AM 细胞具有常驻表型 ( 图 6e) 。
进一步支持 AMs 起源于第 28 天的驻留细胞,向小鼠滴注荧光染料 PKH26 ,这一过程在脓毒症之前标记了几乎所有的 Res-AMs 。在第 28 天,所有的 am 都保持 PKH26+ ,表明它们是感染前或其后代标记的相同细胞 ( 图 6f) 。值得注意的是,观察到在败血症后第 7 天使用 CXCL16 阻断抗体治疗时 AMs 显著降低 ; 然而,在 LLC 发展过程中没有观察到减少。
为了比较 Res-AMs 和 mo-AMs 在脓毒症后减少 T 细胞组织分泌方面的功能作用,测量了氯膦酸钠治疗后 Res-AMs 缺失的脓毒症治愈小鼠或不产生 mo-AMs 的 ccr2 缺陷小鼠中保留的过继转移 T 细胞的数量。研究人员观察到,在 res - am 缺失的败血症治愈受体小鼠中,过继转移的 T 细胞驻留减少,而在 mo- am 缺失的受体小鼠中则没有 ( 图 6g,h) 。最后,研究人员观察到,脓毒症后 28 天, Res-AMs 的消耗增加了脓毒症治愈小鼠的 LLC 表面 ( 图 6i) 。同时,使用 CXCL16 阻断抗体治疗对这种情况下肿瘤生长的影响有限。这一系列结果表明, Res-AMs 是脓毒症治愈条件下诱导 T 细胞居住的趋化因子的主要来源,值得注意的是 CXCL16 ,但其他细胞因子或趋化因子,可能是 CCR2 配体 ( 图 5) ,是 T 细胞充分获得抗肿瘤特性所必需的。
10. 败血症后 T 细胞局部滞留增加
虽然 CXCR6+ 组织驻留 T 细胞的数量在脓毒症后第 7 天达到峰值,然后下降 ( 图 2h) ,但研究人员观察到这些细胞在脓毒症治愈小鼠的肿瘤生长过程中更高 ( 图 3b) 。在免疫再刺激期间,组织内 CXCR6+ T 细胞的积累并不是癌症所特有的,正如研究人员在继发性肺炎期间观察到的类似现象一样,随着组织内 v γ - 6δ T 细胞的逐渐特化而诱导其驻留 ( 图 7a,b) 。
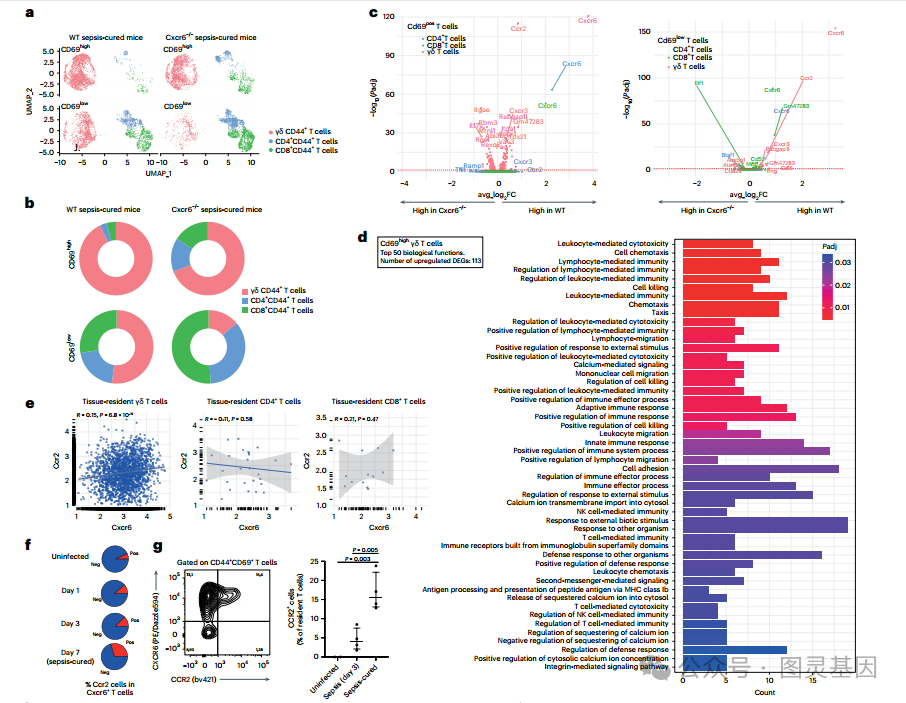
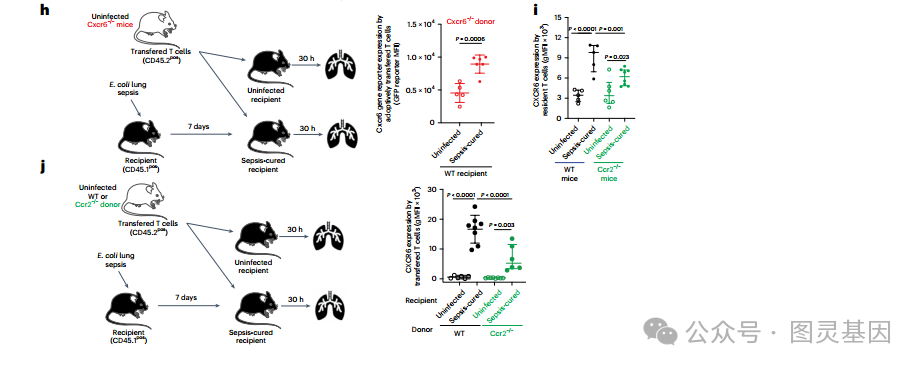
图 4 脓毒症治愈后, CXCR6 和 ccr2 依赖性组织驻留 T 细胞功能的调节
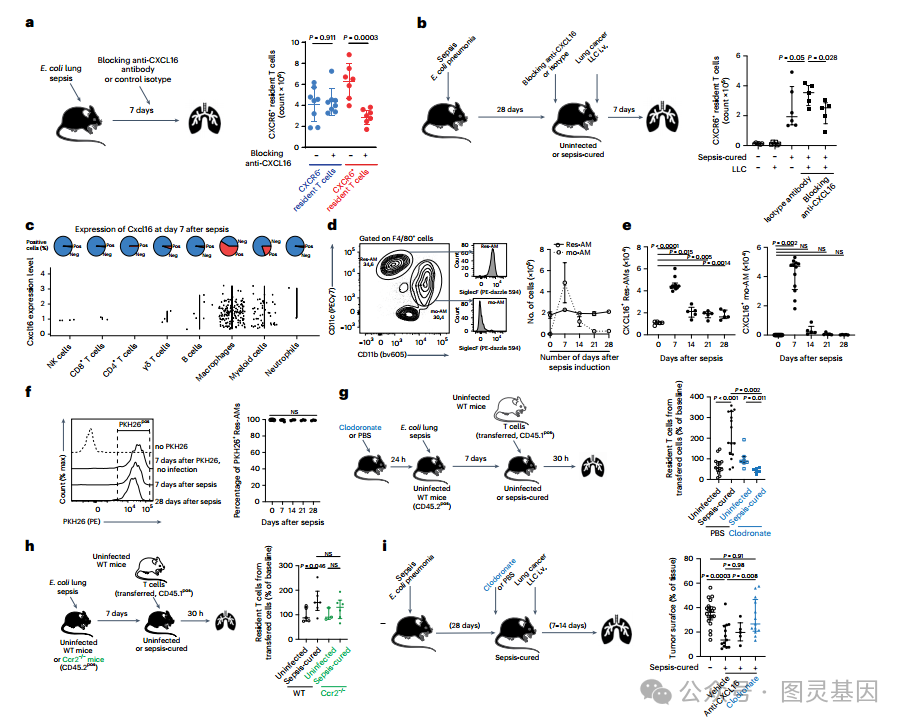
图 5 常驻 am 介导脓毒症后 T 细胞组织输出减少和肿瘤生长
这些发现表明,虽然败血症治愈的组织不能持续维持高数量的 CXCR6+ 组织驻留 T 细胞,但它们在随后的免疫挑战中具有增强的触发能力,以促进 T 细胞驻留。尽管发现在免疫召回期间 CXCR6+ T 细胞的增加需要常驻 am ,但这也可能是由于败血症期间选择的 T 细胞的内在能力。为了测试组织驻留是否是败血症治愈小鼠组织驻留γδ T 细胞中表达的顶级基因之一,在抗原经历的驻留记忆 T 细胞中, CXCR3+ 细胞的百分比高于抗原经历的非驻留记忆 T 细胞,并且抗 CXCR3 已经被证明可以消耗组织驻留 T 细胞。
因此,在败血症后 14 天用抗 cxcr3 抗体处理小鼠,以消耗在第一次败血症期间形成的组织驻留 T 细胞 ( 图 7c) 。观察到抗原经历的非驻留 T 细胞的数量没有减少。小鼠未经治疗 14 天,以形成新的常驻 T 细胞,然后感染大肠杆菌继发性肺炎。研究人员发现,在脓毒症时形成的细胞中,缺失或未缺失的小鼠的常驻 γδ T 细胞总数没有差异 ( 图 7c) 。这一观察结果表明,败血症治愈小鼠的肺组织增加了 T 细胞的驻留,这些 T 细胞在败血症期间没有消耗,但在二次免疫刺激期间被激活。
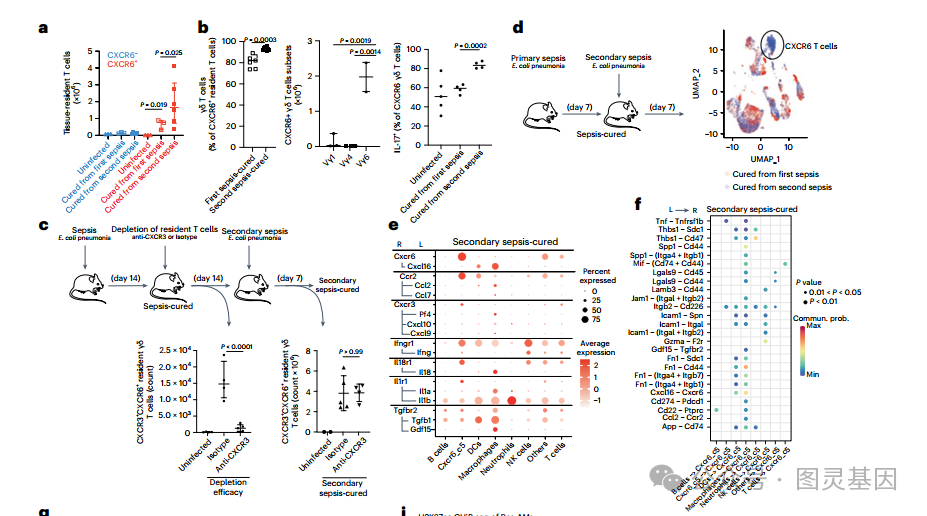
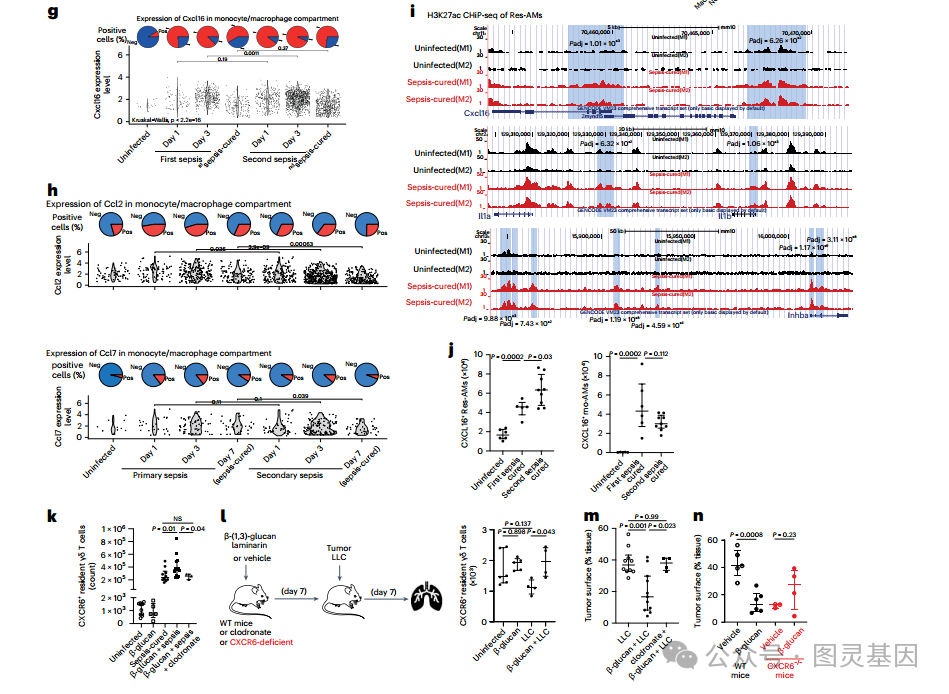
图 6 β-(1,3)- 葡聚糖处理可以模拟训练后 Res-AM 的增加
11.Res-AMs 的表观遗传改变导致趋化因子的产生增加
然后,质疑败血症诱导的 T 细胞生态位修饰是否是一个一体化的、加性的或协同的过程 ( 如果其振幅 ( 细胞因子水平 ) 或多样性 (L-R 对数 ) 在每次败血症后增加 ) 。为了回答这个问题,研究人员通过单细胞 RNA 测序 (scRNA-seq) 表征了继发性大肠杆菌肺脓毒症期间肺组织的免疫细胞组成 ( 图 7d) 。与第一次脓毒症时定义的 R-L 相互作用图谱相比,在第二次脓毒症治愈的小鼠中仍然发现了 Cxcr6-Cxcl16 和 Ccr2-Ccl2 /Ccl7 对,巨噬细胞仍然是潜在 L 的主要来源 ( 图 7e,f) 。值得注意的是,与原发性败血症相比,观察到巨噬细胞在继发性免疫刺激期间产生的趋化因子增加 ( 图 7g,h) 。
研究人员假设这可能与先天免疫训练有关,最近将其定义为先天免疫细胞在第一次激活后对后续刺激的反应增强。训练后的免疫以巨噬细胞的表观遗传调控为特征,之前报道了脓毒症治愈小鼠中 Res-AMs 表观遗传调控的深刻改变。
重新分析这个组蛋白 H3 赖氨酸乙酰化 (H3K27ac) ChIP-seq 数据集,一个与激活和训练免疫相关的染色质标记,在从败血症治愈小鼠收集的 Res-AMs 中发现, H3K27ac 峰在以前开发的 L-R 图谱中的趋化因子和细胞因子的基因组区域增加,如 CxcI16, IL-1a, IL-1b 和 Inhba ,它是 TGFβ 家族的成员 ( 图 7i) 。以及已知参与 T 细胞维持或化学吸引的介质。研究人员通过流式细胞术证实,在继发性脓毒症期间, CXCL16+ Res-AMs 的比例增加,而 mo-AMs 的比例没有增加 ( 图 7j) ,进一步支持脓毒症诱导的免疫训练是由表观遗传改变的常驻巨噬细胞介导的。这一系列分析表明,细胞因子和趋化因子网络多样性在继发性免疫刺激期间没有增加,但每次败血症免疫刺激都会增加巨噬细胞能力产生水平。
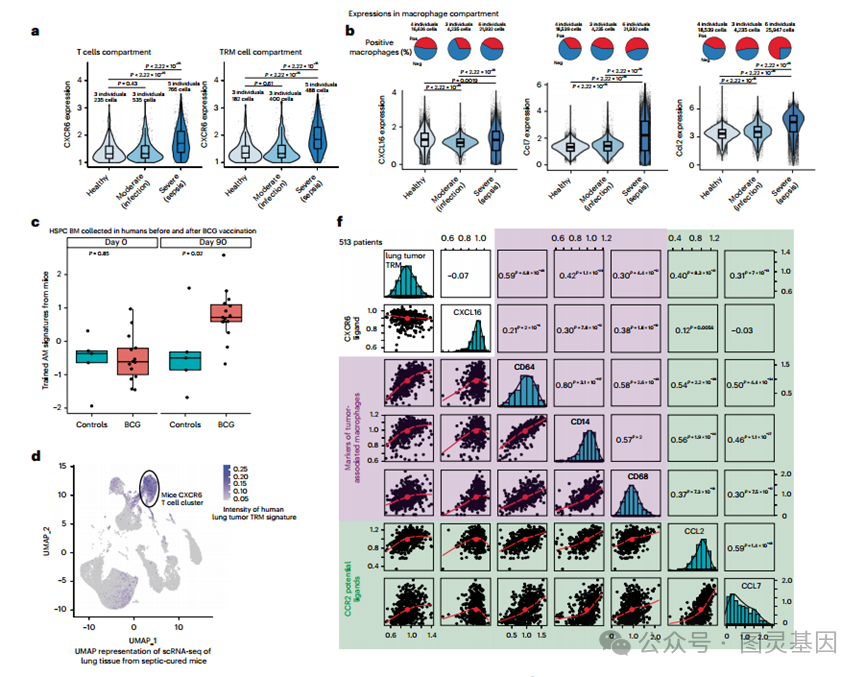
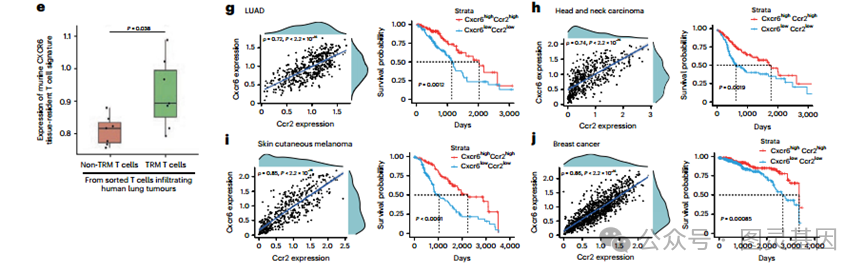
图 7 人类趋化因子巨噬网络和 CXCR6 组织驻留 T 细胞
12.CXCR6+ T 细胞驻留增加是对败血症的特异性反应
研究人员观察到,在金黄色葡萄球菌肺炎治愈的小鼠肺中, CXCR6 组织驻留 T 细胞的数量增加,这支持了该模型是对不同脓毒症原因的共同反应的观点。研究了对金黄色葡萄球菌败血症和 cpgv .15 的反应。研究人员发现,在这种系统性败血症自发治愈的动物的肺、脾、肠或骨髓中, CXCR6+ 组织驻留 T 细胞的数量增加。与脓毒症相比,非脓毒性 SIRS 与人类发生癌症的风险增加有关 ( 图 1c,d) 。因此,假设脓毒症后诱导 CXCR6+T 细胞驻留的细胞因子和趋化因子网络在无菌炎症期间不会发展。
为了验证这一假设,分析了创伤性脑损伤引起的无菌性全身炎症反应小鼠模型中的巨噬细胞和 T 细胞。虽然创伤改变了巨噬细胞的转录组活性,但观察到它们的均匀流形近似和投影 (UMAP) 投影与呼吸道败血症的反应不匹配。值得注意的是,创伤后巨噬细胞腔室中的 Cxcl16 和 Ccl2 表达水平低于大肠杆菌脓毒症后 7 天,表明无菌 SIRS 不会激发脓毒症治愈后描述的趋化因子网络。研究人员还发现无菌 SIRS 后 7 天 CXCR6 组织驻留 T 细胞没有增加。
最后,目标是通过注射β -(1,3)- 葡聚糖来重现败血症诱导的训练免疫,因为这是巨噬细胞表观遗传重编程的一种众所周知的诱导剂。 7 天后,用 β-(1,3)- 葡聚糖的纯化部分 laminarin 治疗或未治疗的小鼠受到大肠杆菌脓毒症或 LLC 的攻击。研究人员观察到,除了 Res-AMs 缺失的小鼠 ( 图 7k, 1) 外,层粘菌素处理增加了脓毒症后 CXCR6+ γδ T 细胞的数量。值得注意的是, Res-AMs 缺失的 laminarin 处理小鼠的存活率下降。最后,重现脓毒症后观察到的结果, laminarin 预处理降低了野生型小鼠的 LLC 生长,但在 res - am 缺失小鼠 ( 图 7m) 或 cxcr6 缺失小鼠 ( 图 7n) 中没有。这一系列实验表明,经过训练的 Res-AMs 对 T 细胞的组织保留在无菌 SIRS 后不会发生,但通过注射 β-(1,3)- 葡聚糖可以部分重现这种现象。
13. 肺脓毒症患者的 CXCR6 T 细胞和训练 AM 发育
采用循环方法,在人体中测试了经过训练的 Res-AMs 诱导 T 细胞驻留的方法。为了实现这一目标,检查了来自中重度 COVID-19 患者支气管肺泡灌洗免疫细胞的公开数据库的 scRNA-seq 数据集。观察到败血症患者中 T 细胞和组织驻留 T 细胞区室中的 Cxcr6 表达水平高于中度感染和健康对照 ( 图 8a) 。研究人员还观察到,脓毒症患者巨噬细胞中 Cxcl16 、 Ccl2 和 Ccl7 的表达水平高于中度感染患者和健康对照 ( 图 8b) ,这表明在人类中,这种免疫现象在脓毒症期间发生,但在中度感染期间没有发生,这与图 1 中描述的脓毒症幸存者与感染幸存者相比癌症风险降低的观察结果一致。在另一个公共数据集中,在接种卡介苗的人的骨髓中发现了训练过的巨噬细胞基因特征,这与研究人员在系统性败血症小鼠模型中观察到的情况相符 ( 图 8c) 。
14.CXCR6+ T 细胞与癌症患者生存的关系
鉴于模拟脓毒症诱导的训练免疫预防和治疗癌症的潜在治疗潜力,研究人员旨在确认训练巨噬细胞来源的细胞因子网络、 CXCR6+ 组织驻留 T 细胞与人类癌症进化之间的关联。首先,对肺癌或结肠腺癌手术患者收集的样本进行免疫组织学分析,发现 CD3 和 CXCR6 共定位。研究人员还检查了从肺肿瘤或真皮中分类的人类组织驻留 T 细胞的公开数据库中的 RNA-seq 数据集。在肿瘤、肺组织和真皮中观察到 Cxcr6 表达水平与组织驻留 T 细胞的基因标记之间存在显著相关性,支持 Cxcr6 在人肺癌组织驻留 T 细胞中的表达。
然后,质疑小鼠模型中描述的 CXCR6 组织驻留 T 细胞群是否可以推断为浸润人肺肿瘤的 T 细胞。为了回答这个问题,从一个公开的数据集中定义了组织驻留 T 细胞浸润人肺肿瘤的基因特征。研究人员发现,在败血症治愈小鼠肺组织 scrna 序列的 UMAP 表示中,该特征的投射与 CXCR6+ T 细胞簇重叠 ( 图 8d) 。研究人员在小鼠数据集上投射人类真皮组织驻留 T 细胞的基因标记时观察到类似的结果。考虑到人类特征在小鼠数据集中投射的偏倚风险,在人类数据集中测试了脓毒症治愈小鼠中定义的组织驻留 T 细胞基因特征的表达水平。
研究人员发现,与非常驻 T 细胞相比,脓毒症治愈小鼠中形成的组织常驻 T 细胞的基因特征在浸润人肺癌的组织常驻记忆 (TRM) T 细胞中丰富 ( 图 8e) 。这些结果支持这样一种观点,即在败血症和诱导训练免疫后积累的 CXCR6+ T 细胞的基因特征接近于浸润人类肿瘤的组织驻留 T 细胞之一。
然后,研究了肺肿瘤 TRM T 细胞特征表达水平与人肺腺癌 (LUAD) 肿瘤 TCGA 数据库中肿瘤相关巨噬细胞标志物 (CD64 、 CD14 和 CD68) 、 Cxcl16 和 CCR2 配体 (Ccl2 和 Ccl7) 的相关性 ( 图 8f) 。肺肿瘤 TRM 特征与检测的巨噬细胞标志物和 CCR2 配体表达水平相关 ( 图 8f) 。这些相关性支持了巨噬细胞释放的趋化因子网络在人肺肿瘤中 CXCR6+ 组织驻留 T 细胞积累中的作用。在 TCGA 数据库的 LUAD RNA-seq 数据集中, CCR2 与 CXCR6 表达水平高度正相关,进一步支持了人类的推断,即 T 细胞上的 CCR2 刺激增加了 T 细胞的 CXCR6 表达 ( 图 8g) 。最后,发现 Cxcr6-Ccr2 共表达水平最高的患者比低表达的患者生存率更高 ( 图 8g) 。
为了评估 Ccr2+ Cxcr6+ T 细胞与人类癌症的一般相关性,将这一分析扩展到几种 TCGA 肿瘤类型,选择那些在法国败血症幸存者队列中风险降低的肿瘤,如宫颈癌和颈癌、皮肤黑色素瘤和乳腺癌。因此,在这些癌症中, Ccr2 与 Cxcr6 表达高度相关, cxcr6high - ccr2high 共表达与生存率增加相关 ( 图 8h-j) 。
15. 讨论
败血症是一种由感染引起的全身性炎症。这种与高死亡风险相关的情况表明,为了根除病原体而产生的免疫反应会对宿主的健康产生负面影响。虽然先天免疫在败血症期间观察到的细胞因子风暴中起着核心作用,但单核吞噬细胞的训练,改变了它们对后续触发因素的反应,可能是个体在整个生命过程中优化免疫病理和耐受之间的利益 - 风险平衡的重要机制。
危险模型理论提出,免疫系统并不区分自身 ( 细胞损伤和癌症 ) 和非自身 ( 败血症 ) ,而是区分可能造成损害的危险和不会造成损害的危险。这一理论的基础是病原体和危险相关的分子模式诱导了一种共同的先天免疫反应模式,反映了每种条件激活的上游受体的重叠 ; 然而,脓毒症后的训练和耐受性都有报道,免疫触发因素的性质和剂量在决定这些相反效应方面起着重要作用。
研究人员发现,与败血症后观察到的相反,无菌组织损伤不会诱导 T 细胞驻留所需的细胞因子和趋化因子网络,并且与患癌症的风险增加有关。这些观察结果完善了危险模型理论,表明自我和非自我触发对免疫系统的教育是不同的,只有病原体相关的分子模式才能增加肿瘤免疫监视。
虽然在哺乳动物中已经证实了训练免疫和感染抵抗的遗传,但骨髓被认为是败血症后单核细胞谱系 ( 单核 / 巨噬细胞 ) 训练的主要部位。呼吸道败血症后, Res-AMs 的训练持续数月,导致早期免疫抑制,表现为低吞噬能力,随后通过增强中性粒细胞对细菌感染产生后期抵抗。尽管一小部分 Res-AMs 来自终生在肺部募集的单核细胞,但这些细胞的更新主要在局部进行,骨髓前体的贡献很小。研究人员认为巨噬细胞对环境的时空适应是一种独立于骨髓祖细胞外周诱导训练先天免疫的体内机制。这种新机制完成了骨髓祖细胞在培养免疫系统中的作用,但并不排除这种作用。为了统一这些观察结果,研究人员提出脓毒症诱导骨髓巨噬细胞的训练,从而增加局部免疫和骨髓祖细胞,从而产生表观遗传重编程的循环单核细胞。
在这里,研究人员证明在外周组织中训练免疫的发展因此表现为固有固有免疫细胞和适应性免疫细胞交叉调节的相互富集和完善。研究人员发现,脓毒症治疗通过增加γδ和 CD4+ CXCR6+ T 细胞在二次免疫攻击期间的组织驻留来降低肿瘤生长,这与先前的研究结果一致,即驻留 γδ T 细胞和 CD4+ T 细胞可以识别恶性细胞,浸润肿瘤并具有细胞毒活性 44; 然而,研究人员观察到,与野生型小鼠相比,未感染的 cxcr6 缺陷小鼠的肿瘤生长下降,并且 Vγ6+ Vδ1+ T 细胞在慢性细菌定植中促进肿瘤细胞增殖,特别是通过延长炎症性肺反应。为了统一这些观察结果,关键是要考虑 T 细胞驻留起始的时间和组织驻留 T 细胞的质量。
关于时间,研究人员证明了败血症训练的巨噬细胞不会诱导慢性组织驻留 T 细胞炎症,而是在继发性免疫挑战时触发这些细胞的驻留。由于慢性炎症是癌症发展的一个危险因素,这是败血症后治愈与细菌定植期间观察到的重要区别。关于组织驻留 T 细胞的质量,发现在脓毒症治愈后,它们从未感染小鼠的 CD8+ 常规 T 细胞转变为 IL-17+ rorr γ T Vγ6δ T 细胞和 CD4+ T 细胞,并且 CXCR6 刺激增加了组织驻留 γδ T 细胞的细胞毒性功能。因此,假设在触发 CXCR6 刺激后,组织驻留 T 细胞在慢性扩张或抗肿瘤的情况下可能是致原性的。
通过补充实验,包括 Res-AM 缺失,趋化因子来源鉴定, CXCL16 阻断, CXCR6- 和 ccr2 缺陷小鼠和 T 细胞过继转移,提供了强有力的证据,证明由训练巨噬细胞产生的趋化因子网络在诱导 T 细胞驻留和控制败血症后肿瘤中的作用。虽然 Res-AMs 的减少减少了脓毒症后组织驻留 T 细胞的数量并增加了肿瘤的生长,但观察到调节单一介质 ( 例如阻断 cxcl16 抗体 ) 并没有显着改变免疫肿瘤控制。这一结果表明,经过训练的 Res-AMs 产生的趋化因子具有互补和潜在冗余的功能。这是针对训练先天免疫的治疗性繁殖的重要信息,支持重编程 Res-AMs 可能是比单一趋化因子调节更有效的策略。
在这种情况下,β - 葡聚糖是一个广泛的 β-d- 葡萄糖多糖家族,其培养先天免疫的能力得到了很好的认可。然而,先前研究中使用的多糖成分的广泛异质性阻碍了纯化β - 葡聚糖作为批准的人类免疫调节药物的发展。研究维持 β- 葡聚糖免疫刺激活性所需的最小链和基序,有助于为癌症治疗药物的批准铺平道路。在这里,使用了一个很好表征的低分子量的棕海藻海藻素的分数。这种 β-(1,3)- 葡聚糖由 ~ 25 个线性葡萄糖基单位 ( 平均分子量为 4 kDa) 和 6 个葡萄糖基单位的短序列组成,这是与 β-(1,3)- 葡聚糖的主要受体 dectin-1 有效相互作用所必需的。
脓毒症在人类中是一种异质性疾病,小鼠模型可能无法再现这种综合征的全部复杂性。研究采用了几种单细菌脓毒症模型,因为它们与许多人类脓毒症病例有关。事实上,大多数败血症病例,如呼吸道感染 ( 结核病、流感和社区获得性感染 ) 、结肠炎和尿路感染,都是由单一细菌病原体引起的,而不是由多种微生物感染引起的。在不同细菌 ( 大肠杆菌和金黄色葡萄球菌 ) 或注射途径 ( 肺或静脉注射 ) 引起的小鼠模型中观察到稳健的结果。
此外,在小鼠模型中观察到的重要机制在人类中也被观察到,包括脓毒症和非感染期间巨噬细胞产生 CXCL16 ,肿瘤组织中 CXCR6+ T 细胞信号与巨噬细胞产生的趋化因子表达水平之间的正相关, Ccr2 和 CXCR6 表达水平之间的相关性,以及在广泛的人类癌症中存活与 Ccr2 - CXCR6 信号之间的关联。这些结果支持小鼠模型和人类数据之间的联系 ; 然而,人体结果支持一种关联,而不是因果关系,因此在实施医疗干预之前需要谨慎。
研究结果可以概括为四个主要发现 :(1) 脓毒症训练的常驻巨噬细胞形成细胞因子和趋化因子网络,增加 CXCR6+ T 细胞的组织驻留,以响应继发性免疫触发 ;(2) 这些组织常驻 CXCR6+ T 细胞在脓毒症治愈后增加抗肿瘤免疫监测 ;(3) 层粘胶蛋白诱导训练免疫可能是一种增加 T 细胞驻留和降低患者肿瘤复发风险的治疗策略 ;(4) 人类肿瘤中的 Ccr2-Cxcr6 特征可以帮助预测生存。这项研究提示了脓毒症长期免疫效应的新机制,将训练的巨噬细胞、 γδ 和 CD4+ T 细胞的组织驻留和癌症免疫监视联系起来。
教授介绍

Antoine Roquilly 教授, 2018 年,被任命为南特大学麻醉学和重症监护正教授,并于 2022 年 1 月成为南特大学 UMR1064CR2TI (前 EA3826 ) Team 6 实验室主任。主要研究方向及成果 : 在肺炎领域开展了互补的临床和基础研究。在医学课程的早期,研究重点是 HAP 的预防、诊断和治疗。自 2021 年起,担任 CIC Pluithématique 的 “ 免疫学和感染 ” 团队主任。在实验室里,研究了肺炎期间炎症对骨髓细胞功能的影响。在墨尔本大学微生物学和免疫学系做博士后期间( 2013-2015 ),研究 SIRS 引起的免疫功能障碍。证明了耐受性 DC 会诱导 CD4 Treg 细胞积累,从而增加对肺部病原体的耐受性( Immunity 2017 )。
参考文献
Broquet, Alexis et al. “Sepsis-trained macrophages promote antitumoraltissue-resident T cells.” Nature immunology vol. 25,5 (2024): 802-819.doi:10.1038/s41590-024-01819-8
链接:http://www.lewenyixue.com/2024/05/31/Nat%20Immu%20%7C%20%E8%84%93%E6%AF%92%E7%97%87%E8%AE%AD%E7%BB%83%E7%9A%84%E5%B7%A8%E5%99%AC%E7%BB%86/

赶快来坐沙发