Cell | 空间多组学分析揭示胶质母细胞瘤的多层组织结构
撰文: huacishu
IF=64.5
推荐度: ⭐⭐⭐⭐⭐
亮点:
本文作者结合了空间转录组学、空间蛋白质组学和计算方法来定义神经胶质瘤细胞状态并揭示其组织结构。这对神经胶质瘤生态系统日益增长的理解增加了空间维度,并可能有助于未来治疗方法的发展 。
以色列魏茨曼科学研究所 Itay Tirosh 教授课题组在国际知名期刊 Cell 在线发表题为 “ Integrative spatial analysis reveals a multi-layeredorganization of glioblastoma ” 的论文。

胶质瘤包含不同状态的恶性细胞。本文作者结合了空间转录组学、空间蛋白质组学和计算方法来定义神经胶质瘤细胞状态并揭示其组织结构。作者发现了三种突出的组织模式。首先,胶质瘤由小的局部环境组成,每个环境通常富含一种主要的细胞状态。第二,特定的状态对在多个尺度上优先接近。这种状态配对在肿瘤中是一致的。第三,这些成对的交互共同定义了一个由五层组成的全局架构。缺氧似乎驱动了这些架构,因为它与包括所有癌症细胞状态的长期组织有关。因此,远离任何缺氧 / 坏死灶的肿瘤区域和缺氧不足的肿瘤区域较少被组织。总之,该研究为神经胶质瘤细胞状态的组织提供了一个概念框架。
1 、胶质瘤的空间转录组学
使用 10X Visium 空间转录组学平台通过 RNA-seq 对胶质瘤进行空间分析(图 1A )。在四个病例中,作者分析了来自同一肿瘤的多个冷冻组织块,每个组织块都从不同的区域分离出来,并注释为坏死、浸润或 T1 对比度增强。分析了 13 个胶质母细胞瘤( GBM )切片和 6 个 IDH 突变的神经胶质瘤切片。这些样本与来自外部 Visium 数据集的 13 个 GBM 切片整合,在质量控制后总共保留了 70618 个点。
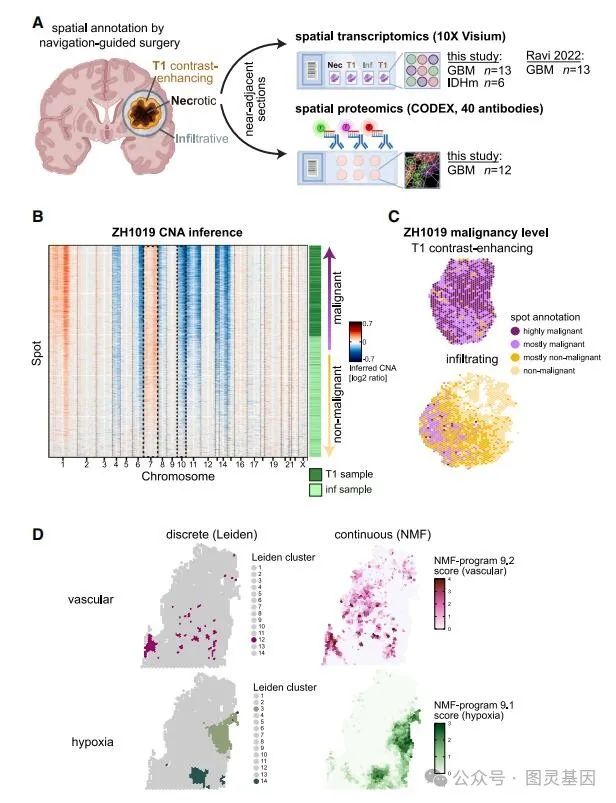
图 1. 实验设计和每个样本的点分类
作者使用了两种最初的方法来对点进行分类和注释。首先,通过平均每个染色体区域的基因表达来进行拷贝数畸变( CNA )推断,将点分为大多数恶性、大多数非恶性或混合型。正如预期的那样,推断的 CNA 包括 GBM ( 7 号染色体增加和 10 号染色体缺失)和少突胶质瘤( 1p/19q 号染色体共缺失)的特征,并与富含癌症的区域(例如,肿瘤核心与浸润区域)显著相关(图 1B 、 1C )。其次,在具有多个切片的病例中,通过每个样本和每个肿瘤的 Leiden 聚类对点进行聚类,并通过基因集富集分析对聚类进行注释(图 1D )。对每个样本进行了非负矩阵分解( NMF ),并导出了在多个参数上一致检测到的稳健表达程序(图 1D )。
2 、胶质瘤中表达异质性的复发模式
为了定义反映表达异质性核心模式的表达程序,作者比较了通过 Leiden 聚类或 NMF 在单个 GBM 样本中鉴定的 492 个基因表达程序。作者发现了相当大的相似性,这使得能够根据它们的基因身份重叠定义 14 组程序,每个程序都涵盖了来自多个 GBM 样本的程序(图 2A 和 2B )。对于每组程序,定义了一个由 50 个基因组成的一致程序,称为元程序( MP ),它反映了 GBM 异质性的复发模式(图 2C )。同样,在 IDH 突变型胶质瘤中定义了 6 个 MP 。
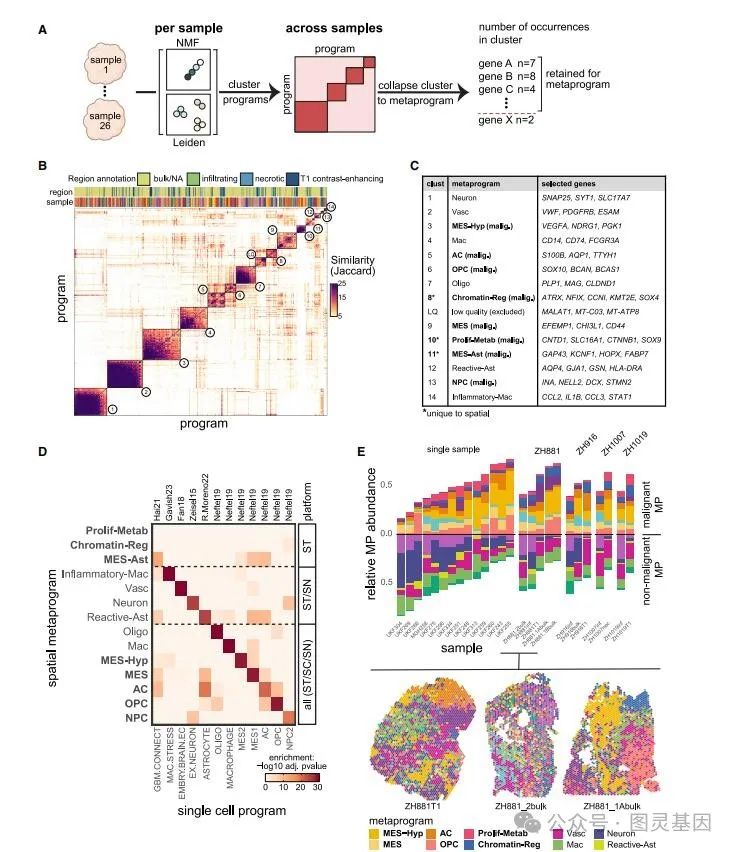
图 2. 导出和注释空间元程序
作者确定了 14 个 GBM 空间 MP ,包括 8 个恶性和 6 个非恶性程序,每个程序都反映了癌症细胞状态或非恶性细胞类型(图 2C 和 2D )。非恶性 MP 包括 Mac (巨噬细胞 / 小胶质细胞)和炎症性 Mac (炎性巨噬细胞 / 中性粒细胞)、 Oligo (少突胶质细胞)、 Vasc (内皮细胞和周细胞)、 Neuron 和反应性 Ast (反应性星形胶质细胞)。在 8 种恶性 MP 中,有 5 种直接映射到单细胞 GBM 状态: MES 缺氧( MES2 )、 MES ( MES1 )、类 NPC 、类 OPC 和类 AC (图 2D )。正如预期的那样,神经发育相关的恶性 MP ( NPC 样、 OPC 样和 AC 样)与相应的非恶性细胞类型特征具有高基因重叠,来源于 scRNA-seq 的相应 MP 也是如此。
接下来,作者通过得分最高的空间 MP 对所有点进行注释。每个 GBM 样本的总体组成高度可变(图 2E ), MES-Hyp 和 Neuron 频率在样本之间变化最大, MES 样频率变化最小。即使是从同一肿瘤的不同区域分离的样本,其细胞类型组成也高度可变(图 2E ),这突出了当单个组织切片被认为代表整个肿瘤的组成时,采样偏差的程度。 MP 对点的分配也与其他与入侵性和连通性相关的特征的分配相关。三种间充质 MP ( MES 样、 MES-Hyp 和 MES Ast )和反应性 Ast 的点都与高连接性有关,而神经发育 MP ( OPC 样、 NPC 样和 AC 样)和细胞周期信号的侵袭性显著富集。
3 、空间蛋白质组学作为胶质瘤定位的补充方法
虽然 Visium 通过覆盖大多数基因提供了全面的数据,但它的空间分辨率较低,不太适合捕捉罕见或空间分散的细胞类型和细胞状态。因此,作者使用 CODEX 作为一种补充方法,在真正的单细胞蛋白质水平上验证他的发现。 40 个标记物抗体组是基于单细胞 GBM MP 和经典标记物设计的,以覆盖几乎所有相关的细胞类型和细胞状态(图 3A 和 3B )。
质量控制后,保留了来自 12 个样品的 428395 个细胞。使用 PhenoGraph 对单细胞的蛋白质表达谱进行聚类,鉴定了所有主要分化的非恶性细胞类型,包括星形胶质细胞、少突胶质细胞、神经元、血管细胞(内皮细胞和周细胞)、 T 细胞、 B 细胞和巨噬细胞 / 小胶质细胞(图 3C 、 3D )。还确定了主要的恶性细胞状态,包括 MES 样、 MES-Hyp 、染色质 -Reg 、 OPC 样、 NPC 样和 AC 样(图 3E )。在 80% 的样本中,位置相邻切片的 CODEX 和 Visium 数据之间的 MP 分配显著相关,这是对 MP 分配的交叉验证(图 3F )。
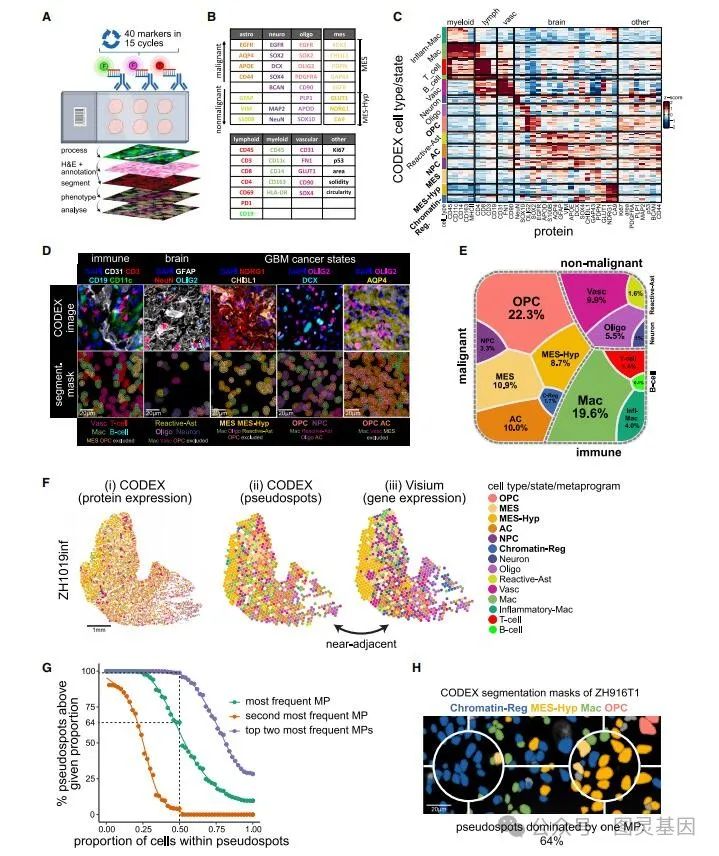
图 3. 胶质瘤的 CODEX 空间特征分析
4 、 GBM 肿瘤包含同时发生的结构化和非结构化区域
在用得分最高的空间 MP 对每个点进行注释后,作者想知道每个样本中的相邻点与分散点在多大程度上富集了每个状态,作者称之为 “ 空间一致性 ” (图 4A )。样本之间的空间一致性各不相同,因此在一些样本中(称为 “ 结构化 ” ),大多数状态往往具有高一致性,而在其他样本中(也称为 “ 无组织 ” ),大部分状态往往具有低一致性(图 4B )。虽然大多数状态的空间一致性根据样本和位置而变化,但少数状态要么一致分组(高空间一致性),要么分散(低空间一致性(图 4C ))。
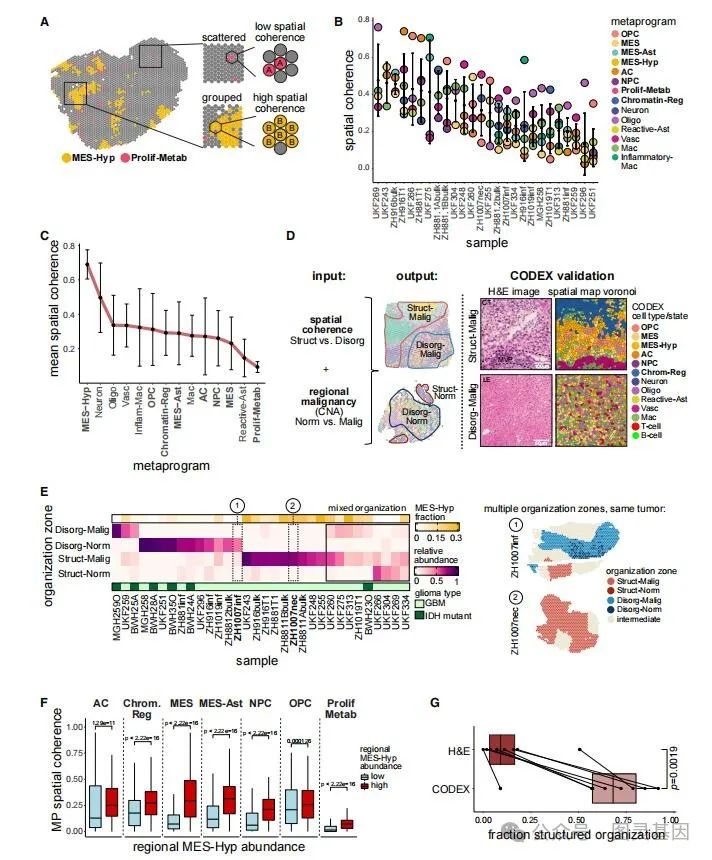
图 4. 细胞状态和组织区域的空间分布
为了进一步探索这种区别,作者设计了一种计算方法,根据其局部环境的一致性,将每个点分类为属于结构化、无组织或中间区域(图 4D )。在肿瘤的核心(富含 MES-Hyp )和正常大脑的浸润区域(富含神经元)都发现了结构化区域(占总点数的 48% )。因此,根据其恶性程度( CNA 信号)进一步细分了结构化和无组织区域,定义了四类: Struct-Malig 、 Disorg-Malig , Struct-Norm 和 Disorg-Norm (图 4D 、 4E )。其余的点被注释为中间点,对应于组织区域之间的边界。所有具有多个切片的 GBM 肿瘤都由结构化和无组织的隔间组成,这表明这些模式的共同出现是 GBM 的一个反复出现的空间特征(图 4E )。 Struct-Norm 区反映了非癌脑实质的组织,神经元的出现频率最高,而 Disorg-Norm 区由癌症浸润程度高的正常脑区组成, Oligo 的出现频率也最高。
5 、癌症细胞状态的空间组织与 MES-Hyp 丰度相关
如上所述, MES-Hyp 在 Struct-Malig 区域富集。即使在从空间一致性的计算中去除 MES-Hyp 点之后,富含 MES-Hyp 的样品也具有高的平均空间一致性。因此,与缺氧相关的组织增加超出了缺氧癌症细胞状态本身,并且与缺乏 MES-Hyp 点的区域中的相同状态相比,靠近 MES-Hyps 点的所有恶性细胞状态都更具组织性(图 4F )。因此,缺氧周围的结构化区域明显比与缺氧结构相关的两种组织学注释 —— 假性栅栏样坏死( PAN )和 MVP 更丰富(图 4G )。因此,缺氧可以提供外力,该外力在组织区域上赋予表达状态的结构化连续性,从而产生通过标准组织学可见的已知局部结构和通过细胞状态的分子谱检测到的先前未被重视的长程结构。 GBM 和 IDH 突变型胶质瘤中的非缺氧区域在很大程度上是无组织的( Disorg-Malig )(图 4E ),与组织学上看到的胶质瘤的整体无组织一致。
6 、成对的状态 - 状态空间关联在样本和尺度上重复出现
为了量化状态之间的空间关系,作者设计了三种互补的测量方法,强调不同分辨率水平下的状态耦合。首先,这两个状态的区域组成,定义为它们在预定义半径(用 r 表示)的六边形窗口中的丰度之间的相关性(图 5A )。第二,两个状态之间的邻接,定义为一个状态在另一个状态的相邻点中的富集。第三,同一点内两个状态的共定位。作者观察到,在越来越大的区域内,状态关系的总体一致性,同时,当将所有措施放在一起考虑时,也确定了特定的尺度依赖性变化(距离依赖性耦合)(图 5B )。
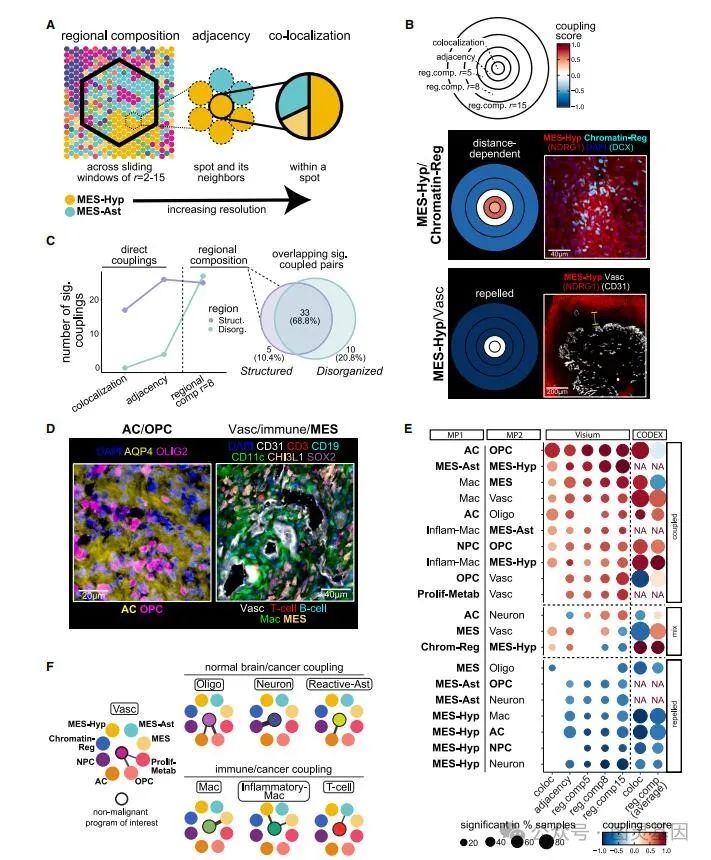
图 5. 跨尺度的状态之间的空间关联
按区域组成来看,结构化和非结构化区域具有相似数量的显著空间关联,这些关联在很大程度上是一致的,而共同定位和邻接措施大多确定了结构化区域特有的相互作用(图 5C )。因此,即使在没有由相邻细胞或点定义的直接空间图案的情况下,区域组成也倾向于保持。
为了确定驱动结构化 GBM 区域组织的最稳健的交互,作者定义了一组状态 - 状态交互的共识集,这些交互由多个度量和多个样本支持。发现结构化 GBM 区域中的一致性相互作用是无组织 GBM 区域的 10 倍(图 5C )。在无组织的 GBM 区域中,唯一一致的相互作用是 MES 样 /Mac 和 NPC 样 /Neuron 。 CODEX 验证了共识交互作用(图 5D )。
对大多数共识相互作用进行了简要总结(图 5E ),恶性状态在很大程度上被共识相互作用分为两组,每一组都有许多组内相互作用:( 1 )神经发育状态( OPC 样、 NPC 样和 AC 样)和( 2 )一组间充质和缺氧相关状态( MES Hyp 、 MES Ast 、 MES 样和染色 -Reg )。神经发育恶性状态不仅相互作用,而且与同一谱系对应的非恶性状态相互作用—— AC 样与反应性 Ast 相互作用, NPC 样与神经元相互作用, OPC 样与 Oligo 相互作用(图 5F )。所有三种间充质状态都与免疫细胞有很强的相互作用,但每种状态都与不同的免疫细胞亚群相互作用(图 5F )。
7 、结构化区域的 GBM 空间组织分层模型
为了检查上述单个交互是否可以结合起来形成更高阶的组织,作者生成了一个网络图,其中节点表示状态,边表示一致交互(图 6A 和 6B )。该图显示了一个五层组织,其中边仅存在于同一层内或相邻层之间。
这种组织似乎由缺氧梯度主导,第一层由 MES-Hyp ( L1 :核心缺氧 / 坏死)组成,第二层由 MES-Ast 、 MES 样和炎症 -Mac ( L2 :缺氧相关)组成。第三层包括可能有助于解决缺氧的免疫和血管生成相关细胞类型和状态,包括 Vasc 、 Mac 和 Prolif-Metab ( L3 :血管生成反应 / 免疫中枢)。 L3 之后,缺氧可能得到解决,从而导致可能更依赖氧气的神经发育恶性状态的存在 ——AC 样、 OPC 样和 NPC 样( L4 :恶性神经发育状态)。最后,非恶性脑细胞类型(反应性 Ast 、 Oligo 和 Neuron )反映了向浸润的脑实质( L5 :脑实质)的转变。将点映射到五层保留了结构化区域和无组织区域之间的区别(图 6C 、 6D )。 CODEX 数据生成了类似的图,其中向 L3 添加了 B 细胞和 T 细胞(血管生成反应 / 免疫中枢)。
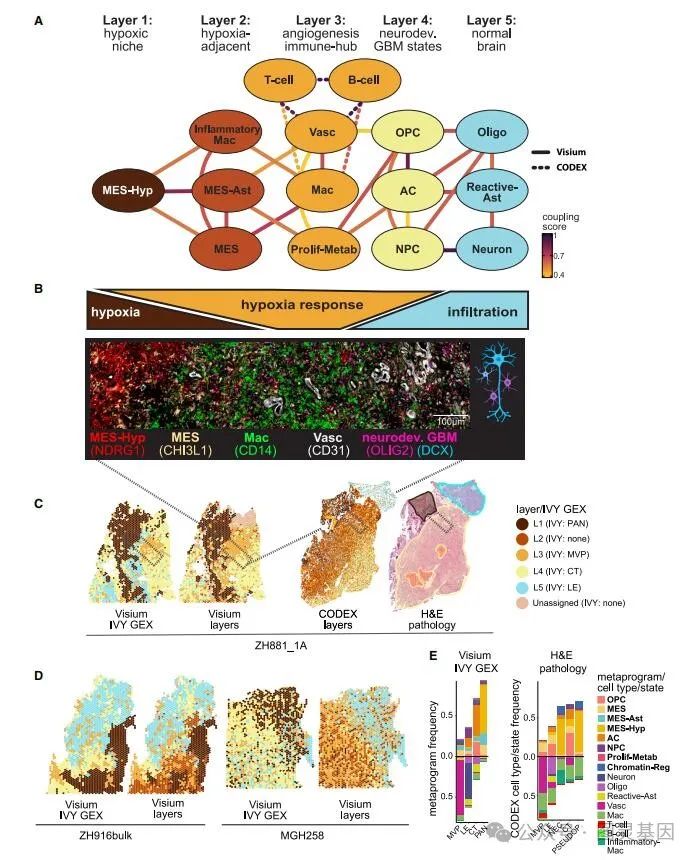
图 6. GBM 空间组织的分层模型
作者发现他们的模型和经典注释之间具有高度一致性, L1 类似于 PAN , L3 类似于 MVP , L4 类似于细胞肿瘤, L5 类似于肿瘤前沿区域。将来源于五层的转录特征与对应于显微解剖的组织病理学特征的转录特征进行比较,显示出除了 L2 之外的总体一致性, L2 没有直接映射到组织病理学特点。因此,通过对基因表达的无监督分析,在很大程度上集中于神经胶质瘤的主要组织学特征,但准确性有所提高,并增加了一层新的缺氧邻近状态( L2 )。此外,现在可以以细胞状态的详细分辨率重新定义经典的组织病理学特征(图 6E )。
8 、结论
总之,作者对神经胶质瘤进行了广泛的空间描述,显示了多个尺度上的空间组织,缺氧作为组织者的突出作用,以及缺乏缺氧的区域的相对紊乱。这对神经胶质瘤生态系统日益增长的理解增加了空间维度,并可能有助于未来治疗方法的发展。
教授介绍

ItayTirosh 教授就职于以色列魏茨曼科学研究所, Tirosh 实验室将计算和实验方法相结合,将人类肿瘤作为一个复杂的生态系统进行研究,在这个生态系统中,不同的癌症和非癌细胞相互作用,共同决定肿瘤生物学和对治疗的反应。利用单细胞技术、计算方法和临床合作来分析人类肿瘤中细胞的多样性。重点是识别重要的肿瘤亚群,如癌症干细胞、耐药细胞、侵袭细胞和对免疫疗法有反应的免疫细胞。然后,研究它们的功能、调节和脆弱性,最终目标是开发更好的癌症治疗方法。
参考文献
Greenwald AC, Darnell NG, Hoefflin R, et al. Integrative spatial analysisreveals a multi-layered organization of glioblastoma. Cell. Published onlineApril 16, 2024. doi:10.1016/j.cell.2024.03.029
链接:http://www.lewenyixue.com/2024/05/14/Cell%20%7C%20%E7%A9%BA%E9%97%B4%E5%A4%9A%E7%BB%84%E5%AD%A6%E5%88%86%E6%9E%90%E6%8F%AD%E7%A4%BA%E8%83%B6%E8%B4%A8%E6%AF%8D%E7%BB%86/