高分综述:肿瘤微环境驱动癌症发展
长久以来,人们认为癌症是由基因突变逐渐积累而成的遗传疾病。但是,让研究人员困惑的是:那些携带致癌突变的胚胎细胞是如何在小鼠身体内和谐发育的呢?又或者,仅仅表达一两个致癌基因的成体细胞怎么就能在某些情况下“变脸”成为恶性肿瘤呢?这说明环境因素可能会点燃表观遗传的“火种”,让细胞的命运发生非遗传性的改变。今天,小编要和大家分享一篇2024年2月发表在Nature Reviews Cancer(影响因子:78.5)上的文章,作者在这篇综述中聚焦于某些癌基因如何在干细胞和周围环境之间开启一场“误会对话”,从而以非遗传的方式扭曲正常组织的发展进程。同时,作者还回顾了这种情况下肿瘤微环境对癌症发生的影响及其在临床上的指导意义。让我们一起揭开这个神秘的癌症发展之谜吧!
一百多年来,癌症一直被描述为一种遗传性疾病。大多数人类癌症包含2-8个驱动突变。例如,在结直肠癌中,进展通常与APC、RAS、SMAD和TP53突变的累积相关。这些研究认为体细胞突变积累达到致癌负荷的阈值是恶性肿瘤状态起始和进展的基础。
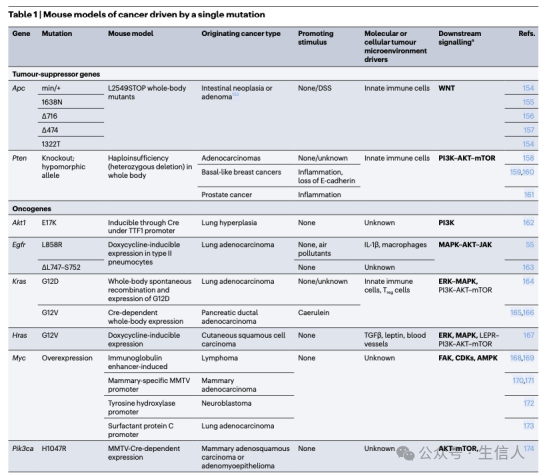
然而现在越来越多对小鼠和人类儿童的研究均表明,少数突变即可诱发具有侵袭性的恶性肿瘤(表1),甚至非转化的上皮细胞也可以自发地在远处器官中转移和存活。事实上,很久以前就有迹象表明,癌症不仅仅是遗传问题。例如恶性畸胎瘤细胞可能有助于嵌合小鼠的健康组织形态发生。存在致癌突变的细胞是否会引发肿瘤还受其他高危因素影响,例如肥胖、饮食、慢性炎症、环境污染、激素变化、组织损伤和许多其他影响生物体健康的畸变,进而影响癌前组织干细胞的微环境。
在过去的十年里,研究人员重点研究肿瘤微环境(TME)各个组成部分的时空变化以及转化细胞在进展为恶性、侵袭性和转移性癌症时的变化。这些研究发现了许多新的癌症驱动因素,包括一些基因改变,例如致癌组蛋白突变,以及许多非基因改变,例如组蛋白和DNA修饰以及肿瘤转录调控的改变,以及TME的细胞外基质(ECM)、组织结构和物理力的改变。此外,以正常组织为对照来看,大多数癌症的TME都很复杂,由多种细胞组成,其中包括适应性和先天免疫细胞、成纤维细胞、血管、淋巴管、神经元和脂肪细胞。此外,肿瘤干细胞(CSC),一种自我更新、维持和传播肿瘤生长的细胞,充当细胞交换机,整合来自TME的各种信号,并传回TME共同促进癌症进展。这些非遗传因素因素是恶性肿瘤形成和进展的关键,且通常是主要驱动因素。因此,作者在本篇综述里介绍了携带单一致癌突变的细胞是如何通过与环境相互作用,驱动表观遗传重编程从而促进肿瘤的发生和发展。此外,作者还讨论了将肿瘤微环境和表观遗传驱动因素作为癌症治疗靶点的潜力。
TME包括免疫细胞、CAF、血液和淋巴管系统和神经,以及ECM、基底膜、生长因子和代谢物等细胞外成分,所有这些都已知会影响肿瘤的发生和进展。
哺乳动物上皮内的细胞竞争是组织形态发生过程中发生的自然生理过程,表现为健康的细胞识别并杀死不健康的邻近细胞,然后将其从组织中清除。因此肿瘤的形成要求转化细胞超越健康的细胞,获得增殖优势,以避免细胞死亡和清除。
最近在乳腺类器官中研究了人类癌症肿瘤发生中的细胞竞争,其中转化的细胞只有在被转化的邻居包围时才能有效增殖。因此,为了使转化细胞的克隆出现,它们必须产生足够的旁分泌信号以阻止健康邻居的竞争。事实上,在肠隐窝中,携带KRAS或PI3K突变的转化肠细胞将骨形态发生蛋白配体分泌到邻近的健康隐窝中,以抑制正常干细胞活性从而获得竞争优势。
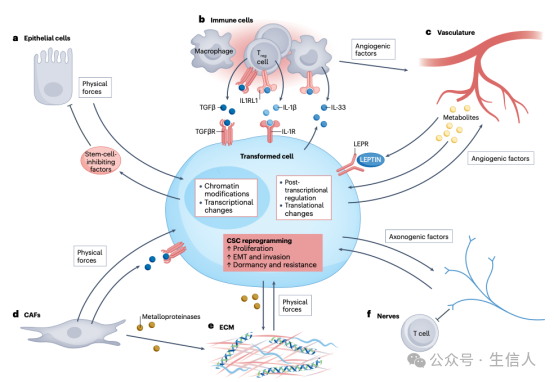
这些研究表明从肿瘤发生的早期阶段开始,在没有基因突变积累的情况下,转化的上皮细胞可以(并且必须)改变其周围的上皮,以成功进展为恶性肿瘤(图1)。然而在某些情况下,特定突变(例如TP53和NOTCH1)可以在皮肤和食道中长期积累表型,而不会发展为恶性肿瘤或被相邻突变消除,这表明除了致癌突变之外,肿瘤发生还需要生态位重塑。
图1. 转化细胞和微环境之间的串扰诱导癌症干细胞的形成
炎症是跨组织癌症发生和进展的关键驱动因素。炎症和癌症之间的第一个联系来自经典的皮肤化学致癌研究,其中用经常诱发Hras突变和乳头状瘤的强效诱变剂DMBA处理小鼠,然后再用炎症刺激物TPA处理小鼠可加速小鼠发展为恶性鳞状细胞癌。同样,接触胰腺炎诱导剂雨蓝素会使胰腺导管细胞中携带单一致癌 Kras G12D突变的小鼠更容易进展为导管腺癌。这些例子表明,尽管长期以来认为必须发生累积的自发遗传突变事件才能发展成恶性肿瘤,但在最初的遗传损伤之前或之后发生的炎症刺激可以取代这种需要。
炎症和癌症之间关系的另一个有趣的发现是暴露于炎症刺激物导致的皮肤干细胞的表观遗传记忆可以在炎症消退且恢复正常后持续很长时间。这些“记忆域”嵌入在一组关键炎症基因的增强子中,使它们的基因保持开放状态,在遇到压力时可以快速、超敏感地重新激活。
CAF是具有肿瘤异质性表型的TME细胞,在ECM沉积和重塑中发挥关键作用,并且在与癌细胞和免疫浸润白细胞的通讯中发挥重要作用。最早的一项研究发现在基质成纤维细胞中敲除TGFβ的共受体基因Tgfbr2时,小鼠出现了自发的上皮前胃肿瘤。此后的研究发现,CAF足以通过转化的上皮细胞驱动前列腺中的肿瘤发生,从而减少了癌症中高突变的需要。进一步的,CAF对上皮肿瘤发生的强大作用也已在遗传背景正常的成纤维细胞中得到证实。此外,研究表明CAF能够改变TME的代谢和缺氧环境,从而改变CSC生态位。
尽管基质和转化上皮细胞之间的许多相互作用受到分泌的细胞因子和生长因子的影响,但其他相互作用可能是由细胞-细胞和细胞-ECM相互作用的快速变化或致癌突变引起的异常机械力触发的,包括基质ECM和基底膜的硬度,其中大部分基质ECM是由CAF分泌的I型胶原蛋白负责的。
TME对癌症另一种影响是血管生成和淋巴管生成,这两者都是由来自CSC的信号触发的。著名的基因是编码血管内皮生长因子A(VEGFA),它是肿瘤相关血管生成的有效诱导剂,可由早期致瘤微环境中的多种因素诱导,包括综合应激反应、缺氧和 TGFβ信号传导。此外,最近的研究已经开始探索神经在癌症进展中的影响,然而研究没有一致的促进或抑制肿瘤的作用。总之,即使相关的CSC具有较低的致突变负荷,脉管系统在促肿瘤方面也具有强大的力量。
TME和肿瘤细胞之间的通讯塑造了癌症表观基因组,其中包括DNA、组蛋白和其他染色质相关蛋白的翻译后修饰。这些改变可以留下持久的标记,影响CSC 的转录和翻译。尽管它们经常对细胞行为产生类似的结果,但表观遗传修饰与基因突变不同,因为它们具有潜在可逆性,这使得它成为未来开发抗癌疗法的一个有趣领域。
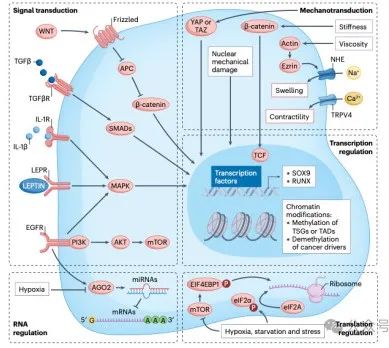
TME 触发的促肿瘤信号传导通常会导致转化细胞中的转录因子失调。MAPK通路被酪氨酸激酶受体(如EGFR)激活,进而激活RAS和/或RAF,改变转录因子(如JUN、FOS、ELK、ETS、MYC、MSK,甚至还有β-连环蛋白,WNT信号传导的淋巴增强子结合因子(LEF1,也称为TCF1)辅助因子。尽管该通路上的基因在癌症中经常发生突变,但该通路可能会被TME过度激活(图 2)。
随着肿瘤发生,复杂的外部因素和下游信号转导通路的相互作用可以深刻改变 CSC的染色质组织。最近的高通量技术能够绘制可及染色质区域、转录因子的结合、染色质修饰和三级染色质动力学图谱,开始揭示染色质的非遗传变化如何促进肿瘤的发生和进化。一种有意思的新发现是肿瘤与其微环境之间的动态串扰产生选择性压力,从而触发组织中通常不活跃的转录因子。
TME的代谢状态也会影响癌症表观基因组学。例如,许多DNA和组蛋白去甲基酶的活性依赖于辅助因子α-酮戊二酸,因此受到TME中α-酮戊二酸与琥珀酸比率的影响。
此外,TME介导的三级染色质拓扑变化也会促进癌症的进展。例如,某些癌症中发生的DNA甲基化表观遗传增加可以掩盖CCCTC 结合因子(CTCF)结合位点,重塑染色质的三维拓扑结构,从而使有效的增强子与致瘤基因的启动子接触。然而,该领域仍处于起步阶段。
调节转录稳定性的机制(例如miRNA)和翻译也被认为是表观遗传调节剂。在癌症中,来自TME的外部信号可以干扰这些生物学过程,从而促进转化、肿瘤发生和进展。
许多miRNA与癌症相关,它们的失调可能是由突变介导的,但TME在调节这些转录后调节因子的表达中也发挥着关键作用。例如,缺氧和EGFR信号传导(后者因突变以及 TME 细胞分泌的生长因子而过度激活)调节 argonaute 2 (AGO2) 蛋白的磷酸化,从而调节 miRNA 的成熟,提示了miRNA机制作为经典致癌突变的肿瘤驱动替代方案的重要性(图 2)。
癌细胞必须制定策略,使其能够在压力条件下生存,同时抑制抗肿瘤免疫反应。不利的TME,例如营养剥夺、氧气限制、高代谢需求和氧化应激,都会引发内质网 (ER) 应激的细胞状态。通过激活内质网应激传感器和适应性应激反应,癌细胞可以抵御环境攻击,增强其转移和耐药能力。这些反应的核心是转化机制。通过抑制全局翻译,同时允许一些关键的应激反应mRNA的翻译,癌细胞可以更好地应对环境压力。其中涉及两个关键目标。一个是RAS-MAPK和PI3K-mTOR信号级联对真核翻译起始因子4B(eIF4B) 的磷酸化限制了帽依赖性翻译;另一个是eIF2α的磷酸化,由四种应激激酶中的任何一种介导。其中一种受氨基酸剥夺调节,另一种受 DNA 损伤调节(图2)。
CSC与其TME之间复杂的通讯在癌症治疗中发挥着重要作用。免疫疗法的最新进展清楚地表明了这一点。例如,虽然来自TME的信号可以增加CSC抵抗细胞毒性T细胞免疫治疗的能力,但免疫治疗本身可以通过刺激细胞毒性T细胞分泌干扰素来促进肿瘤干性,干扰素反过来激活CSC中的促转移转录本。
鉴于TME的异质性及其对抗癌治疗的动态适应,针对TME的干预策略也具有挑战性。保持健康饮食、定期锻炼和低压力已被证明可以改善治疗反应。其中涉及的关键分子是LEPR和瘦素,二者的高水平通常会加速癌症进展。但肥胖与癌症之间的联系不仅仅限于此,脂质和其他因子(例如脂因子和全身炎症)的许多组织变化可能会进一步加剧癌症的发展。
总体而言,西方世界饮食的后果及其引发炎症反应的能力可能是广泛且有害的,不仅因为炎症和癌症之间已确定的联系,而且还因为组织干细胞具有持久的炎症经历的表观遗传记忆,这一记忆甚至可以遗传给后代。最后,干细胞经历的表观遗传记忆不仅限于炎症,还包括迁移记忆和干细胞可塑性。总之,干细胞(可能还有其他长寿细胞)保存过去经历记忆的能力可能会深刻影响小鼠和人类对许多癌症的易感性。
表观遗传记忆领域的一个令人困惑的问题是它如何能够传播这么长时间。尽管迄今为止的注意力集中在组蛋白修饰上,但对DNA甲基化的影响可能会产生更长期的影响。
事实上,我们现在知道许多案例,其中关键的促癌抑癌基因可以通过表观遗传手段而不是突变来调节,从而达到与基因突变产生的结果相似的结果。例如,在散发性乳腺肿瘤中,CpG甲基化介导的乳腺癌易感基因BRCA1的抑制赋予了“BRCAness”肿瘤表型,类似于 BRCA1 突变携带者中常见的情况。此外,作为p53活性的重要调节剂,SRC可能磷酸化p53的抑制剂MDM2来抑制p53肿瘤抑制功能。这些情况下的患者可能表现出与基因突变患者类似的临床表型。然而,这些人的筛查和治疗目前仍然是未知领域。
影响癌症表观遗传学的宿主因素的整体系统性也很重要。除了TME,影响全身健康的因素,包括饮食、压力、睡眠和激素变化,也会影响肿瘤进展和对治疗的反应。例如,情绪困扰已被证明与黑色素瘤患者对免疫检查点阻断的不良反应相关。此外,小鼠和人类的昼夜节律影响乳腺癌的转移潜力,大多数转移播种发生在睡眠期间。最后,几十年来人们已经认识到接受激素替代疗法的绝经后妇女患癌症的风险会增加。
本综述重点关注肿瘤生物学的非遗传驱动因素。其中许多驱动因素很久以前就被发现了,但在基因组革命和癌症遗传学出现的最初几十年里相对被忽视了。只有当人们越来越清楚地认识到突变负担并不能解释癌症的许多特征时,癌症遗传学家才开始将注意力转向非遗传变异来寻求解释。复杂的分子工具和高通量技术的出现,与突变负荷保持较低且恒定的动物模型相结合,极大地提高了我们对 TME及其对癌症进展重要性的认识。
在过去的十年中,新的数据表明,对于大多数癌症来说,一旦干细胞发生最初的致癌突变,非遗传驱动因素可能会导致许多推动肿瘤发生和进展的因素。人类癌症研究表明,遗传多样性是肿瘤进展中相对较早的事件,然而伴随着肿瘤发生和转移的遗传改变很少。因此,仅将癌症视为一种遗传疾病是明显不够的。
组织干细胞和TME之间复杂的通讯回路的分子机制仍在揭开,这些非遗传因素在驱动肿瘤进展中的贡献对于新疗法的开发至关重要,如前所述的压力暴露后的持久表观遗传记忆,DNA的甲基化记忆。作者在这篇综述中强调,在许多情况下,突变累积可能会不可逆转地锁定表观遗传途径,这些途径会通过CSC及其周围TME之间的不适应性串扰而被激活。
过去几十年的重点是癌症的基因组测序,未来十年可能会重点关注通过癌症的空间高通量配体-受体分析来了解细胞通讯网络。先进的计算工具现在使得整合表观遗传学观点成为可能,并且人类类器官培养和小鼠遗传学的进步也有助于推动相关研究的发展。关于改变癌症表观基因组的肿瘤结构驱动的机械力的研究也取得了进展。
总之,肿瘤发生和发生的过程可以被视为遗传和非遗传因素之间的动态平衡,这些因素是为适应特定的癌细胞起源及其在肿瘤进展过程中不断变化的微环境而定制的(图3)。
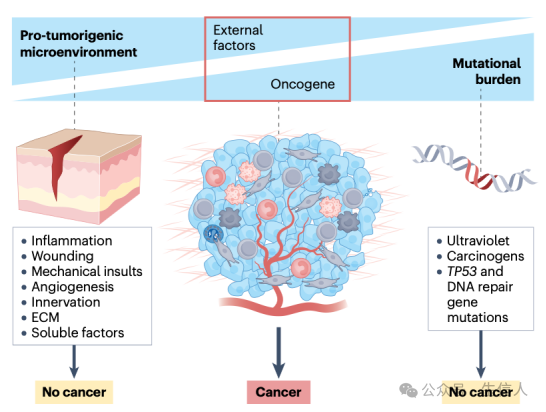
图3. 突变与肿瘤微环境之间的平衡驱动肿瘤的发生和进展
版权声明:本文为“乐问号”作者或机构在乐问医学上传并发布,仅代表该作者或机构观点,不代表乐问医学的观点或立场,不能作为个体诊疗依据,如有不适,请结合自身情况寻求医生的针对性治疗。
链接:http://www.lewenyixue.com/2024/04/26/%E9%AB%98%E5%88%86%E7%BB%BC%E8%BF%B0%EF%BC%9A%E8%82%BF%E7%98%A4%E5%BE%AE%E7%8E%AF%E5%A2%83%E9%A9%B1%E5%8A%A8%E7%99%8C%E7%97%87%E5%8F%91%E5%B1%95/
THE END